

The Watchmaker argument, refuted by evolution?
This video is a powerful tool to demonstrate why micro does not lead to macroevolution, why Darwins Theory of evolution does not withstand scrutiny and the enormous biological
challenge that would have had to be overcome to go from unicellular to multicellular life.
https://www.youtube.com/watch?v=MdjjqBI_EzI
Has Paley's Watchmaker argument been debunked?
https://reasonandscience.catsboard.com/t2860-has-paley-s-watchmaker-argument-been-debunked
Some requirements for transition to multicellularity: 10.00
1, Cell differentiation: 13.32
2. Morphogenesis 18.15
3. Defining each Cell's specific function 25.40
4. Cell migration 27.07
5. Connecting adjacent cells 31.53
6. Cell-Cell communication 33.24
7. Adaptation & microevolution 43.35
8. The genetic program limiting body growth 47.12
9. Gene regulatory network 50.28
10. Setting up sexual reproduction 58.34
Endnotes: 1.02.54

William Paley was an English Christian apologist, and philosopher. He is best known for his natural theology exposition of the teleological argument for the existence of God which made use of the watchmaker analogy.
He believed that the complexity and perfection of nature were powerful arguments for the existence and attributes of God.

Paley famously made an argument by analogy of organisms as watches and of God as a watchmaker, according to which complex structures, like a watch, could not emerge accidentally but require the existence of a rational designer, in this case a watchmaker

Paleys watchmaker analogy is a classic: Paley writes: In crossing a heath, suppose I pitched my foot against a stone, and were asked how the stone came to be there, I might possibly answer, that, for any thing I knew to the contrary, it had lain there for ever: nor would it perhaps be very easy to shew the absurdity of this answer.

But suppose I had found a watch* upon the ground, and it should be enquired how the watch happened to be in that place, I should hardly think of the answer which I had before given, that, for any thing I knew, the watch might have always been there.

Yet why should not this answer serve for the watch, as well as for the stone? Why is it not as admissible in the second case, as in the first? For this reason, and for no other, that, when we come to inspect the watch, we perceive (what we could not discover in the stone) that its several parts are framed and put together for a purpose,
that they are so formed and adjusted as to produce motion, and that motion so regulated as to point out the hour of the day; that, if the several parts had been differently shaped from what they are, of a different size from what they are, or placed after any other manner, or in any other order, than that in which they are placed, either no motion at all would have been carried on in the machine, or none which would have answered the use, that is now served by it.

Without knowing about biology as we do today, Paley made observations, which are spot on, and have astounding significance and correctness, applied to the reality of molecular biochemistry. Let's list the points he mentioned again. If the parts are differently shaped, differently sized, placed after any other manner, or in any other order no motion would be the result.

the individual parts of the watch must be designed in order to be shaped and sized correctly, so they require an intelligent designer.

Each part must be complementary in order to be able to be placed at the right place and the assembly also requires the right sequence. The same is the case in a multicellular organism. This principle is called structural complementarity and is the very essence of biomolecular recognition.
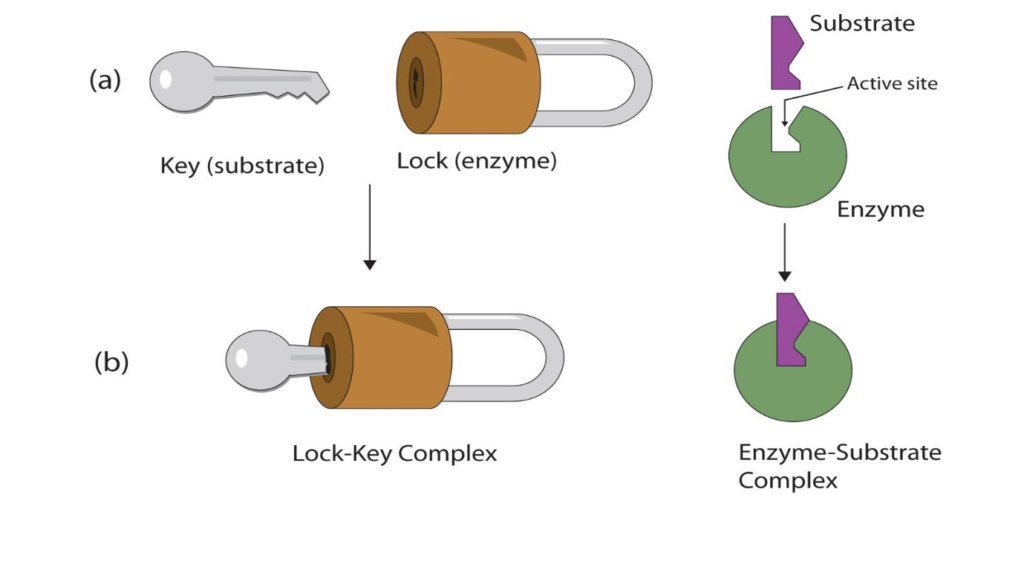
In the same sense as in a watch, one part like an apoprotein without its substrate or prosthetic group must precisely fit and be able to interact with other parts. In biology, structural complementarity is the significant clue to understand the functional properties of biological systems.
Biological systems, from the macromolecular level to the cellular level, operate via specific molecular recognition mechanisms based on structural complementarity: A sperm, for example, recognizes an egg. All these interactions involve structural complementarity between molecules.
So according to the modern evolutionary claims, the watchmaker argument has been refuted by the theory of evolution.

Richard Dawkins borrowed the name from Paley for the title of his 1986 book The Blind Watchmaker, which attacked the idea that complex life needed a religious explanation. And still, today, when the watchmaker analogy is invoked, the immediate answer is: The analogy has been refuted through evolution by natural selection.
But has it? The emergence of multicellularity was ostensibly a major evolutionary leap. Indeed, most biologists consider it one of the most significant transitions in the evolutionary history of Earth’s inhabitants.
We will address this question by giving at what hurdles unicellular organisms would have had to overcome in order to evolve into multicellular organisms.
How do changes in development create new body architectures? Are heritable changes enough, given the constraints imposed by the necessity of the organism to survive as it develops? These questions are critical in molecular
biology, physiology, cell biology, genetics, anatomy, cancer research, neurobiology, immunology, ecology, and evolutionary biology.
The study of development has become essential for understanding all other areas of biology. In turn, the many advances of molecular biology have finally given elucidating answers. Have they been predicted correctly and in a satisfactory manner by Darwin's theory of evolution? The advance of science has it made possible to solve the riddle.
The black box has been opened. Has the evidence led to confirm evolutionary predictions, or have they have been falsified? Evolution would have had to go in a gradual slow, increasing manner from one eukaryotic cell to an organism with two cells, and so on, to get in the end an organism with millions, billions, or even trillions of precisely differentiated cells like the human body with 37 trillion cells.
Let's suppose there were unicellular organisms, eukaryotic so-called protozoans, and evolutionary pressure to go from one to two cells. What and how many mutations would be required in the genome and have to be selected and fixed? Mutations would have to provide the selection of a considerable number of new internal cell functions and created NEW information for these to emerge.
There would have to be set up a way to divide the tasks and assignments of cooperation to consume energy, survive, and reproduce. But how could just this apparently tiny, but in reality huge step in the molecular world have occurred? And why should it have? How did cells supposedly make the transition from life as single cells to split tasks, set up the information to start existing as bicellular organisms,

and associate and cooperate as teams such that they work as a single, cohesive unit? The change would englobe the modification of shape, divide functions and assign to each cell special tasks, structure, and entirely new features of development, ways to connect and stay connected to each other, communicate, and cooperate in order to start doing things in a combined manner.
And all this transition would have to be advantageous during each transitional step, resulting in an organism having an architecture where specialization of each cell in their functions would provide an advantage of survival. If they would duplicate and stick together, without specializing, they would just remain a group of cells or a colony.
Evolutionary literature claims that such transition supposedly occurred 25 times independently, which is called convergent evolution since cell-to-cell adhesion or communication is achieved in various ways in plants, animals, fungi, and algae.
1. Set up cell type and phenotype of the new cells
2. Set up morphology, size and shape of each new cell, and how its new agglomeration leads to new functional structures like tissues and organs organized in a non-random, functional manner.
3. Defining each cell's specific function after differentiation
4. Cell migration, set up and define the position and place in the body of each cell type.
5. Set up specific cell adhesion mechanisms for each cell type
6. Set up Cell-Cell communication channels and signalling networks
7. Set up mechanisms for adaptation and homeostasis specifically for the new emerging multicellular organism
8. The genetic program limiting body growth and cell cycle counter
9. Set up of the gene regulatory network orchestrating gene expression
10. Setting up sexual reproduction

As evidence reveals, a considerable number of innovations would have to be implemented, as listed above. The technical demands of multicellularity are extraordinary. Cells that commit to living together need a whole new set of tools. They have to come up with ways of sticking together, communicating, and sharing oxygen and food.
They also need a master developmental program, a way to direct specific cells to take on specialized jobs in different parts of the body. And a self-destruction program is necessary. The process is called apoptosis. The genes participating in adhesion in animals might have been extant in the unicellular ancestor of animals.
Most of the domains typically found in animals are present for example in choanoflagellates. However, the function of such a diverse set of adhesion molecules in a unicellular organism is not yet known. There are more demands and requirements which will not be elucidated in this video.
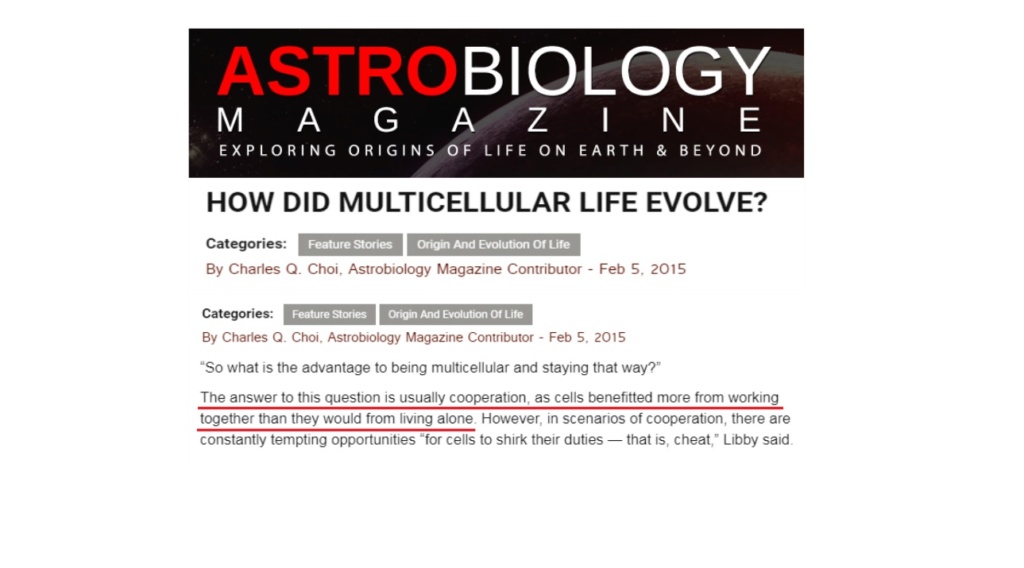
Why did cells not remain single-celled? In Astrobiology magazine, the answer to this question is usually cooperation, as cells benefitted more from working together than they would from living alone. Hummm. Really ??!! An article in Science magazine in June 2018 admitted that the transition to multicellular life as momentous as any in the history of life, until recently we had no idea how it happened.
The gulf between unicellular and multicellular life seems almost unbridgeable, so they confessed. If so, does the explanation above not seem, let's say, a little bit simplistic and naive? or, worse, even pseudo-scientific?

In the paper: The Origins of Multicellularity, the author writes: My line of reasoning is that the first step in the evolution of multicellularity was a size increase due to an accident, in other words, a mutation that prevents the daughter cells from separating. If the larger cell mass has any advantages, such as ensuring the safety of the germline by producing a protected internal environment, then natural selection will see to it that the novelty is retained.
Cells require ultrasophisticated proteins that keep them together. Arguing that an accident did hold them somehow together seems nothing more than just so guesswork. We will give now a closer look at the mentioned innovations.
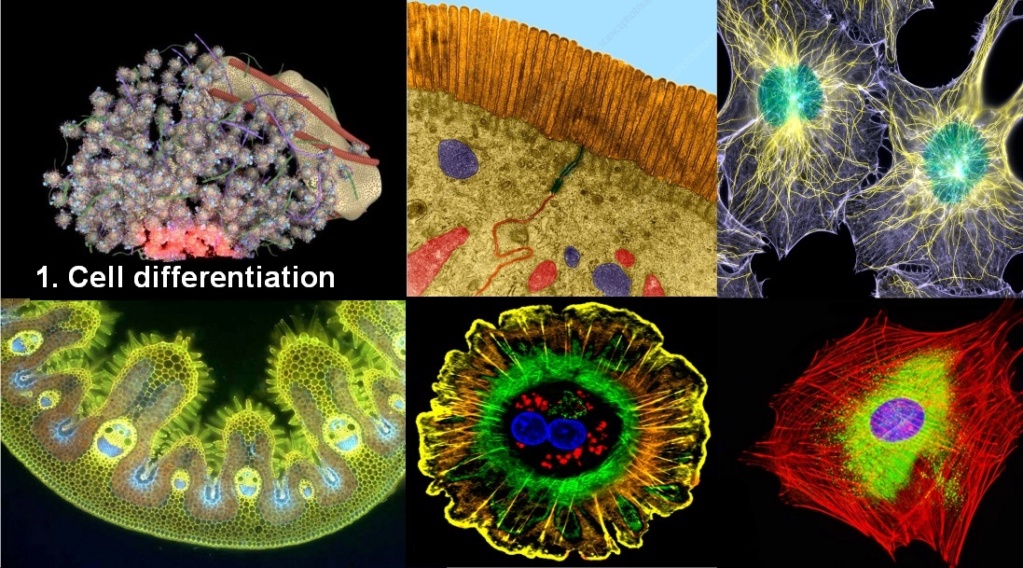
Cell differentiation. That is already a huge venture. Stem cells can generate transit cells and, finally, differentiated cells. In most cases, stem cells can produce more than one type of differentiated progeny (multi- or totipotent). So the transition from a Cell containing just the information to make one cell, to a cell with two or more differentiated Cells would have to occur, sharing tasks.
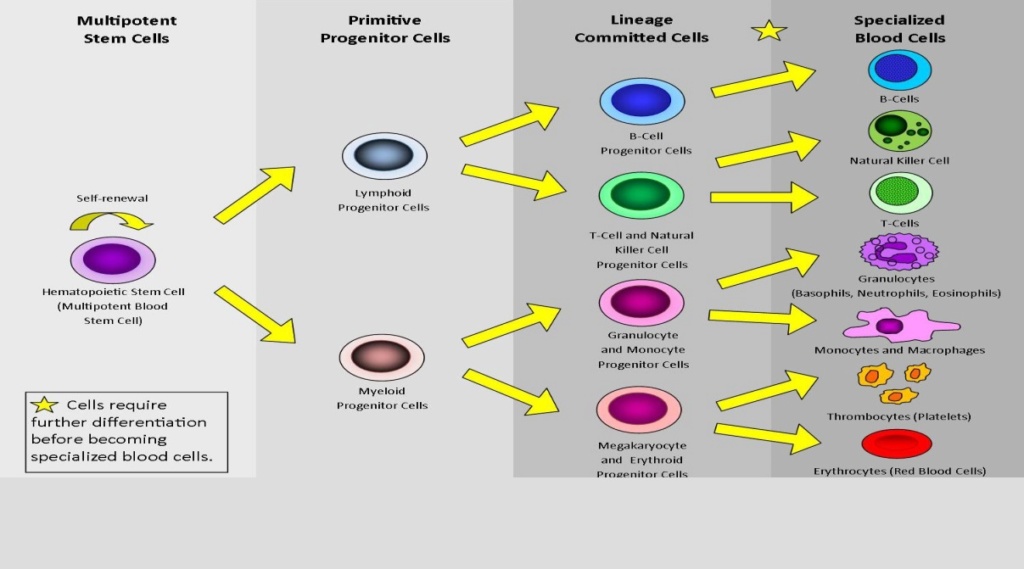
Now let us suppose early earth was populated by unicellular life forms, like protozoans. Then the transformation to multicellular organisms began to evolve. Pluripotent cells would have had to become able to produce specialized cells, where each one of them would have had function only together with all other cells, like ten different types of cells forming blood.
This differentiation had to be organized and coordinated in advance on a systems level above the individual cell level. Foresight is required to know the end function of blood in a joint venture of all required individual blood cell types.
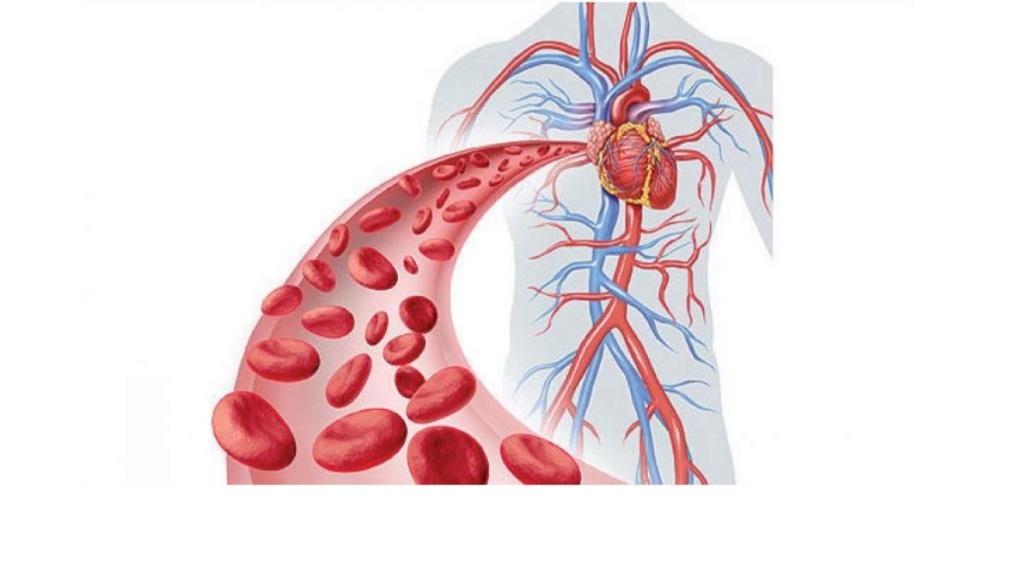
Blood is only functional if it carries all ten different cell types, which have each different, vital functions for life. Platelets, for example, are essential components of blood whose function is to stop bleeding by clumping and clotting blood vessel injuries.
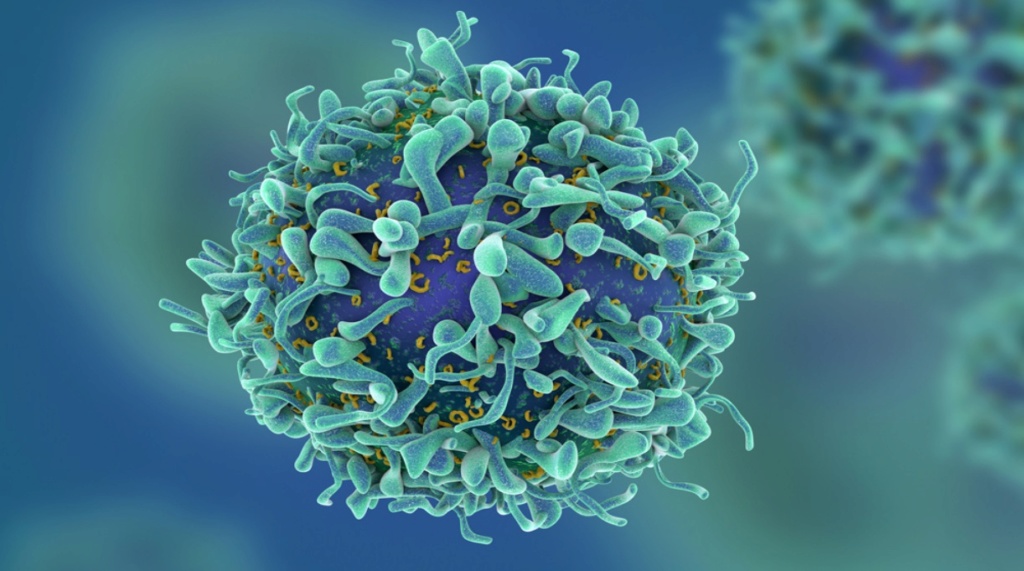
T cells are essential for human immunity. They orchestrate an immune response and play important roles in all arms of immunity.

Red blood cells are the most common type of blood cells and essential for the vertebrate organism's delivering oxygen to the body tissues—via blood flow through the circulatory system.

At least six of the different cell types in the blood are life essential. If one is missing, life using cardiovascular systems is not possible. Had these blood types not to emerge together, to make the life of a multicellular organism like vertebrates possible?
Moreover, Vascular cells have also been shown to be part of the niche that controls the maintenance and differentiation of hematopoietic stem cells which form all blood cells and reside in the bone marrow. This forms an entire interdependent system where bones had to be existent, providing bone marrow, the place where blood and vascular formation takes place.
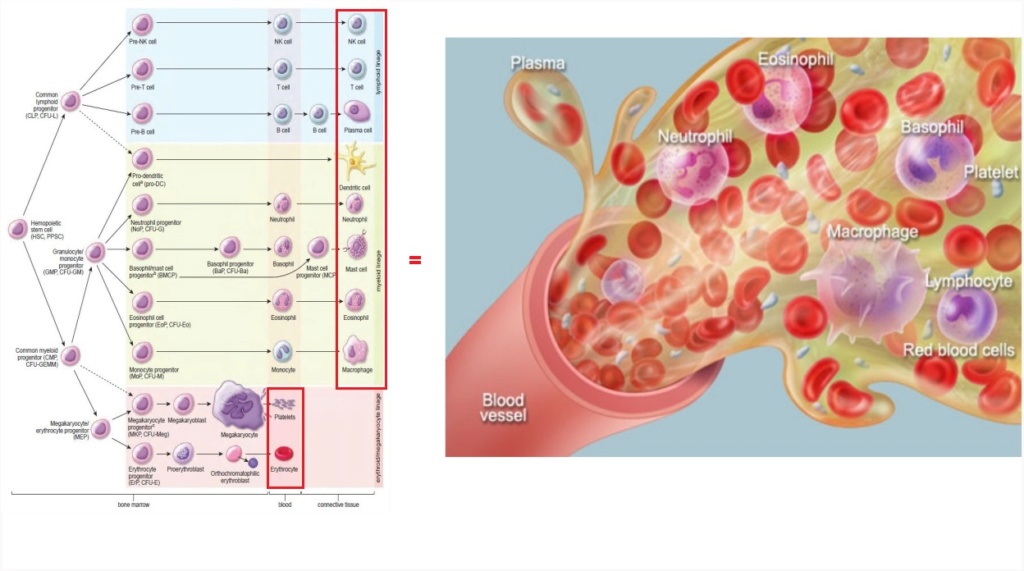
Most blood types require intermediate steps in their development until becoming final cell types used in blood. These intermediate cell types have no function in the organism. Why would evolution produce them or natural selection select them? We are talking about an enormous number of mutations required to produce each of the ten different blood cells.
Many cells play many roles during development, going through stages that are no longer seen in adulthood. How can natural selection account for the emergence of these cells? The role of some cell types is to activate specific genes in nearby cells, and once they accomplish their task, they die. This is clear evidence of an orchestrated, pre-programmed process.

I think Byron Bledsoe brings it pretty well to the point in this quote. Irreducible complexity is powerful evidence of creation, and the vascular system composed of veins and blood cells, and bone marrow where hematopoietic stem cells are produced, is an interdependent system, and so a very illustrative example. But how does science explain the origin of the blood vascular system?

It is evident that the authors in the rather recent paper cited above from 2013, Evolutionary origins of the blood vascular system and endothelium, evolution is assumed, but the authors have nothing to offer besides declarations of vague guesses and assertions based on evolutionary beliefism.
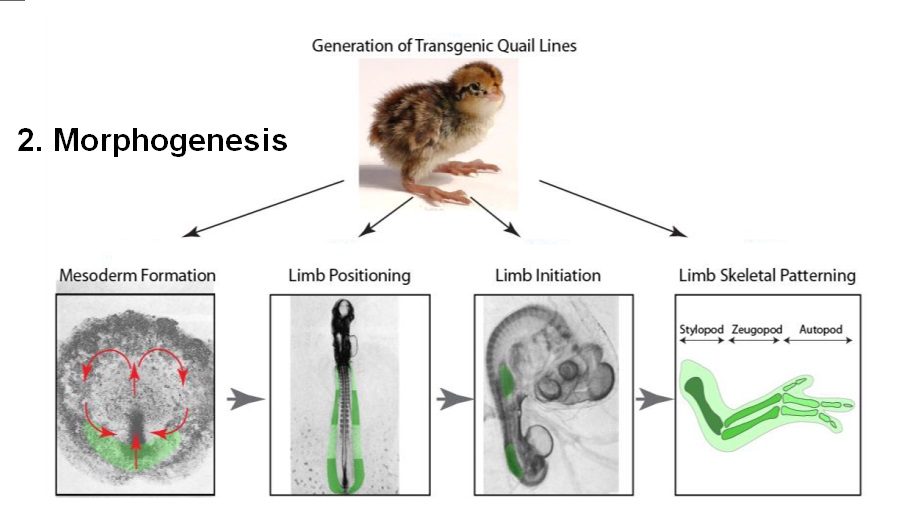
Morphogenesis

Morphogenesis depends on a serious of factors, amongst it defining the size and shape of each cell, which has to organize into functional structures. Differentiated cells are not randomly distributed. Rather, they are organized into intricate tissues and organs. During development, cells divide, migrate, and die; tissues fold and separate.
Genetic and epigenetic information has to be set up to define morphology, function, size and shape of each new cell, and how its new agglomeration leads to new functional structures like tissues and organs organized in a non-random, functional manner.
Cells come in all sorts of forms and sizes, and that is defined based on the functions they will exercise.
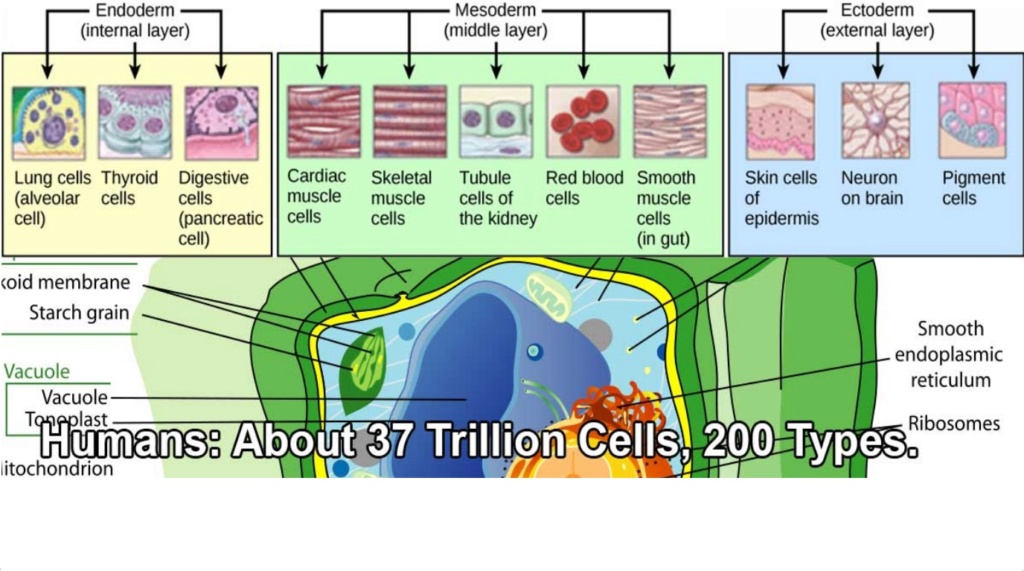
In an article in the magazine wired, HOW LIFE MADE THE LEAP FROM SINGLE CELLS TO MULTICELLULAR ANIMALS, the authors write: After the appearance of microbes, life got more complicated. Cells organized themselves into new three-dimensional structures. They began to divide up the labour of life so that some tissues were in charge of moving around, while others managed eating and digesting.
They developed new ways for cells to communicate and share resources. These complex multicellular creatures were the first animals, and they were a major success. Soon afterwards, roughly 540 million years ago, animal life erupted, diversifying into a kaleidoscope of forms in what’s known as the Cambrian explosion.
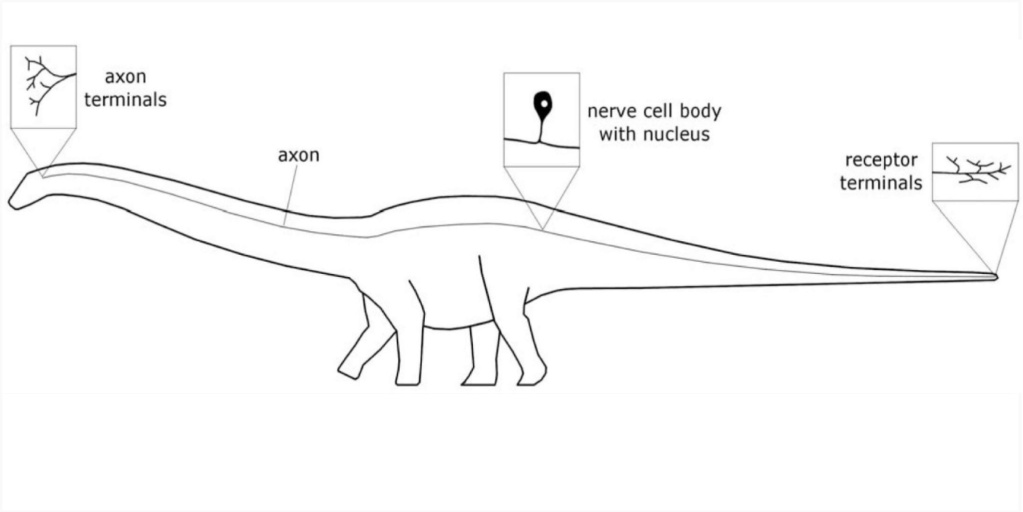
The longest-necked animals of all time were the extinct sauropod dinosaurs, some of which had necks 14 meters long. In these animals, the neurons that comprised the recurrent laryngeal nerve were at least 28 meters long. In the longest sauropods, these neurons may have been 40–50 meters long, probably the longest cells in the history of life.
They were sensory neurons that connected receptors in the skin of the extremities with interneurons in the brainstem, a pattern of neural architecture that is present in all extant vertebrates.
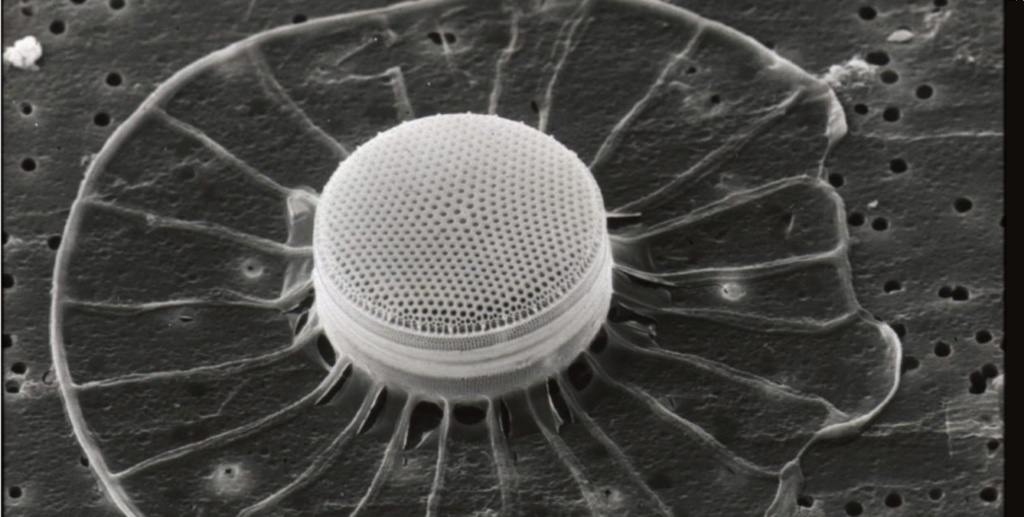
The biggest single-celled organism can be up to 20cm across – Syringammina fragilissima’s one cell branches out into a network of tubes extending over ten centimetres ( or 3.9 inches). As it grows, the deep sea-dwelling creature oozes slime onto the sediment, solidifying its structure.
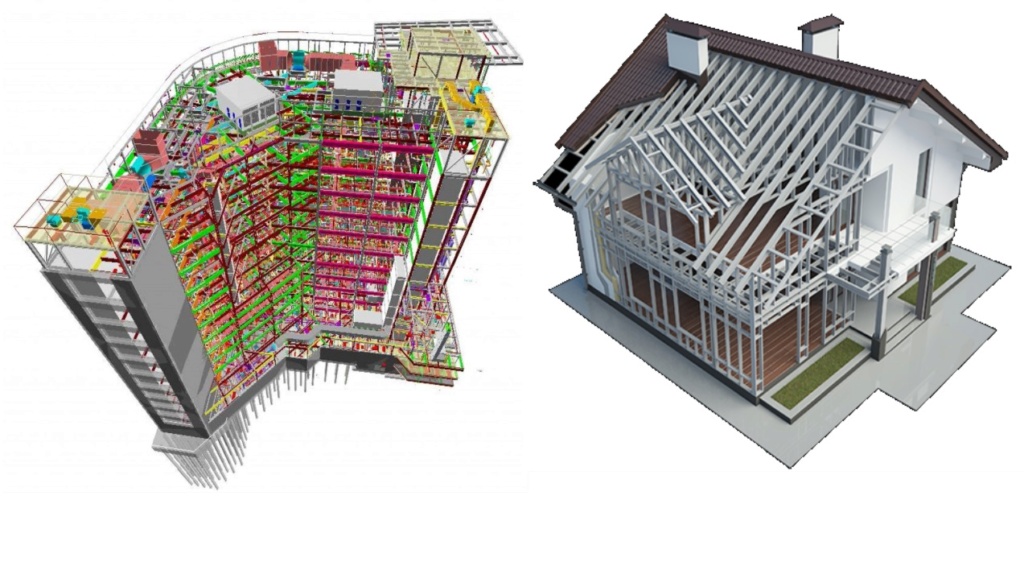
The size and form of a hospital are totally different than of a residential house. What defines its structure, is the scaffold structure, which has to be defined precisely taking the specific demands in consideration.
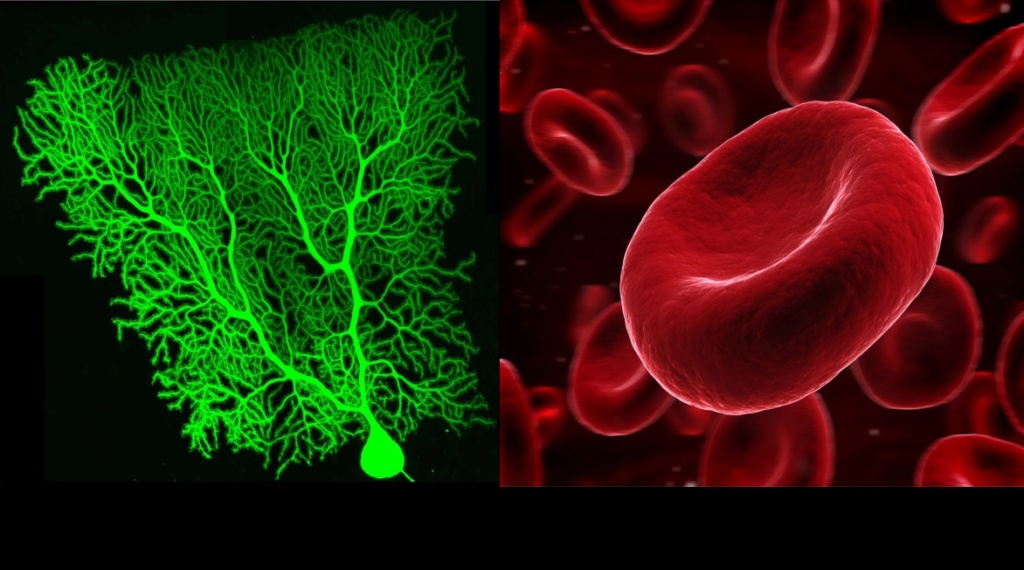
Shape and form of cells have to be defined in accordance with the function that each cell type will perform. A neuron requires a totally different form and shape than a red blood cell.
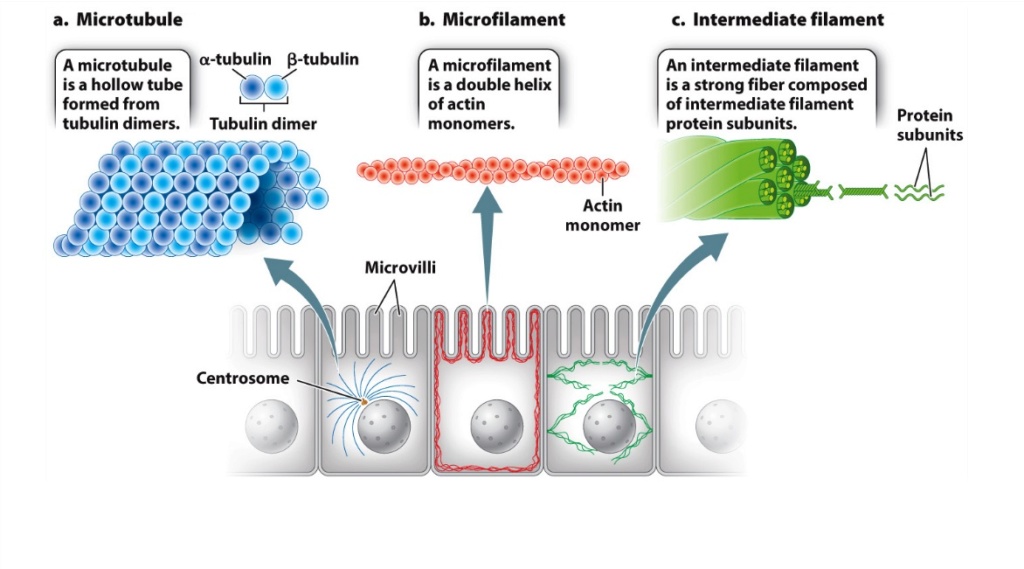
In cells, actin, actomyosin and microtubules that form the cytoskeleton exercise the same function as the scaffold structure of a house or a large building, defining shape and form.
For cells to function properly, they must organize themselves and interact mechanically with each other and with their environment. They have to be correctly shaped, physically robust, and properly structured internally.
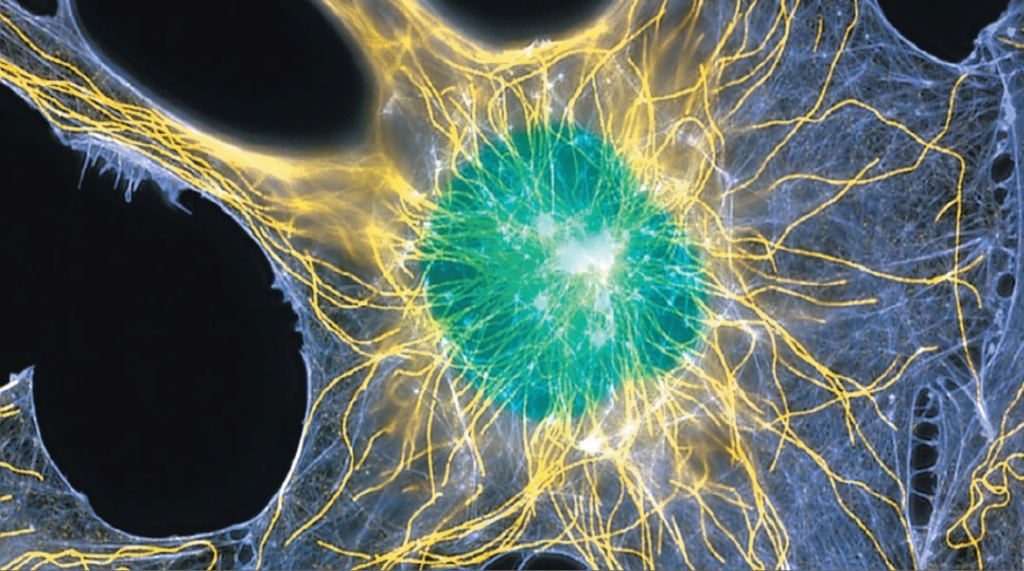
Many have to change their shape and move from place to place. All cells have to be able to rearrange their internal components as they grow, divide, and adapt to changing circumstances. These spatial and mechanical functions depend on this remarkable system of filaments.
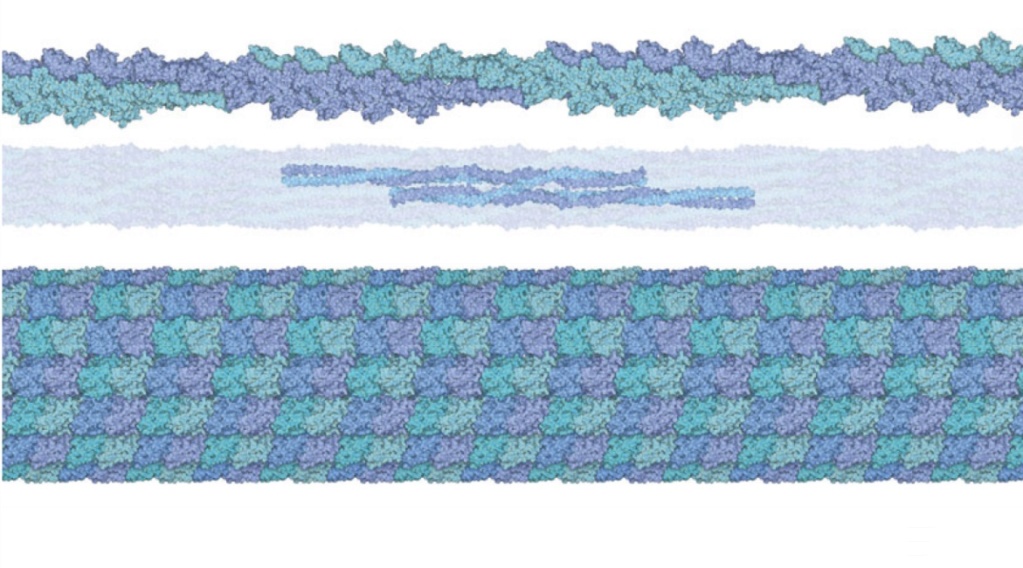
The cytoskeleton’s varied functions depend on the behaviour of three families of protein filaments—actin filaments, microtubules, and intermediate filaments. Microtubules are very important in a number of cellular processes. They are involved in maintaining the structure of the cell and provide a platform for intracellular macromolecular assemblies through dynein and kinesin motors.
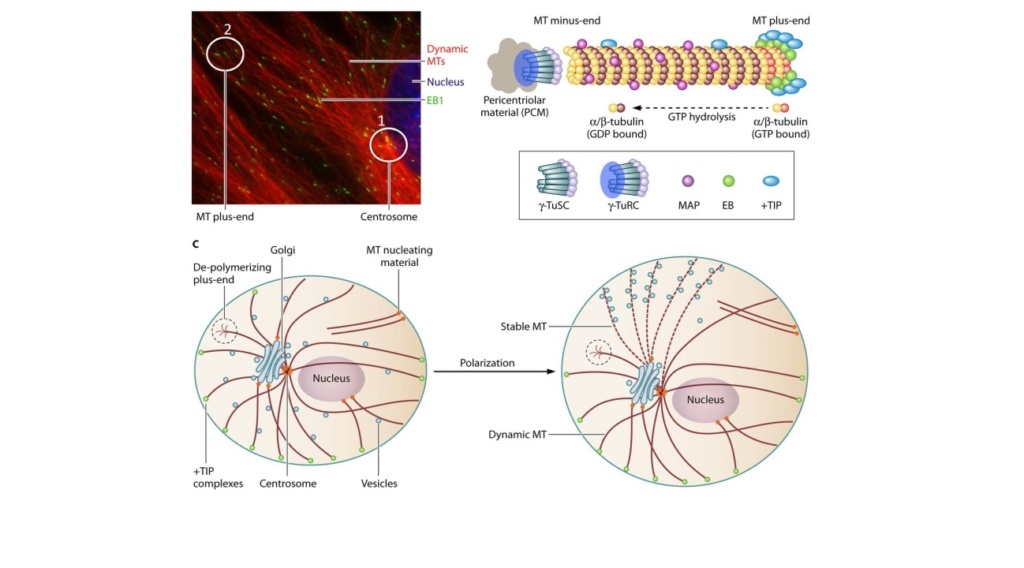
They are also involved in chromosome separation (mitosis and meiosis) and are the major constituents of mitotic spindles, which are used to pull apart eukaryotic chromosomes. The dynamic cytoskeletal and membrane system that governs morphological patterning in cells is fundamental.
This system enables eukaryotes to package signalling molecules in endosomes, tiny vacuoles that bud from surface membranes, and transport them through the cell by means of molecular motors at rates much faster than would be possible by diffusion alone
As a result, eukaryotic cells can change shape or physiology in response to molecular signals, permitting processes such as locomotion of amoebas, and permanent cell differentiation. The dynamic cytoskeleton and membrane system of eukaryotic cells open up possibilities of size, structure, function, and development not available to prokaryotic organisms.
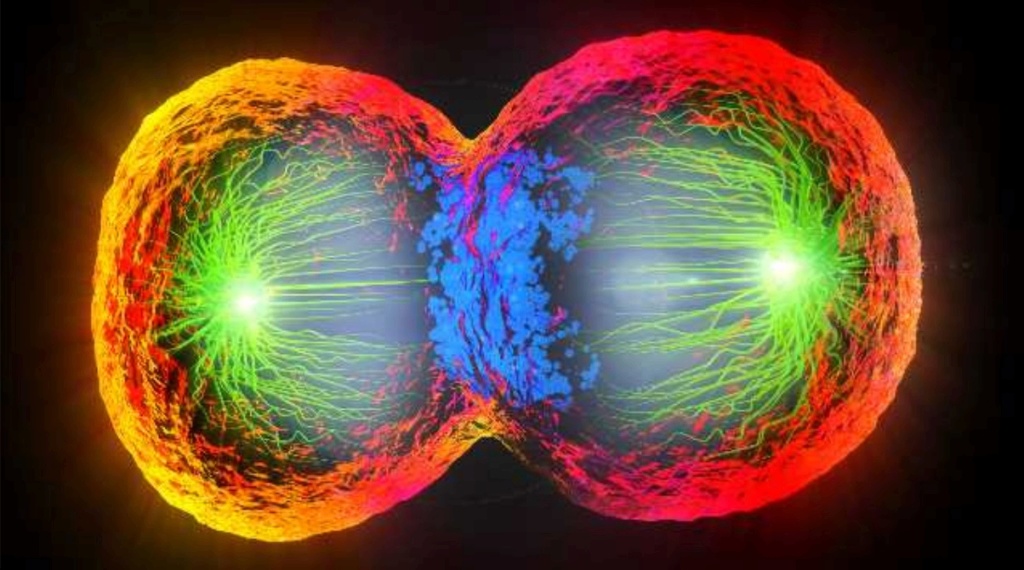
Mitotic cell division is the most fundamental task of all living cells. Cells have the intricate and tightly regulated machinery to ensure that mitosis occurs with appropriate frequency and high fidelity. If someone wants to explain the origin of eukaryotic cells, the arise of mitosis and its mechanism and involved cell organelles and proteins must be elucidated.
The centrosome plays a crucial role: it functions as the major microtubule-organizing center and plays a vital role in guiding chromosome segregation during mitosis. These architecturally perfect structures are essential in many animal cells and plants.
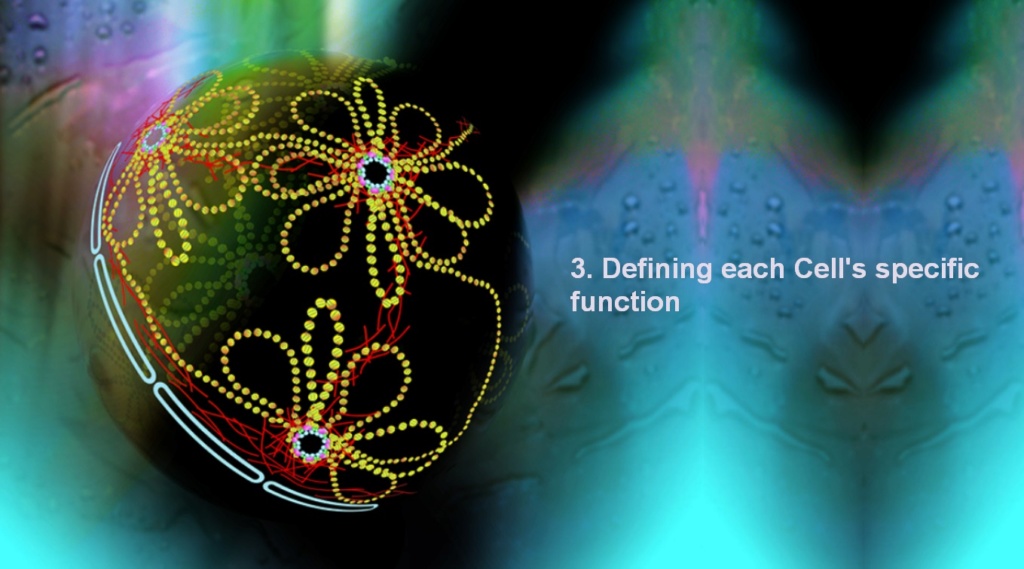
Defining each Cell's specific function Genes define the amino acid sequence that makes proteins, and specific sets of proteins define tissue-type. There are no proteins for hemoglobin in neuronal cells and there aren't proteins that make voltage-gated ion channels in red blood cells.
These functional elements of the cells, along with other cell-specific proteins, are responsible for making a neuron a neuron and a red blood cell a red blood cell. Yet, both cell-types have the genes for each of these cell-specific proteins, in fact every eukaryotic cell with a nucleus in a multicellular organism contains a full set of all tissue-specific genes as part of the full genome.
Tissue-specificity must then arise from how genes are differentially expressed, meaning that in some cell lineages certain sets of genes are silenced, while in others they are expressed – leading one line of progenitor stemcells to differentiate into neurons and another to differentiate into erythrocytes, or any of the other approximately 200 different cell types of the human body.

Another issue that has to be explained is individual, and a more advanced stage, collective cell migration which has a key role during morphogenesis and during wound healing and tissue renewal in the adult. It is a central process in the development and maintenance of multicellular organisms.
In addition to displaying a coordinated migratory behaviour, collectively migrating cells move more efficiently than if they migrated separately, which indicates that a cellular interplay occurs during collective cell migration.
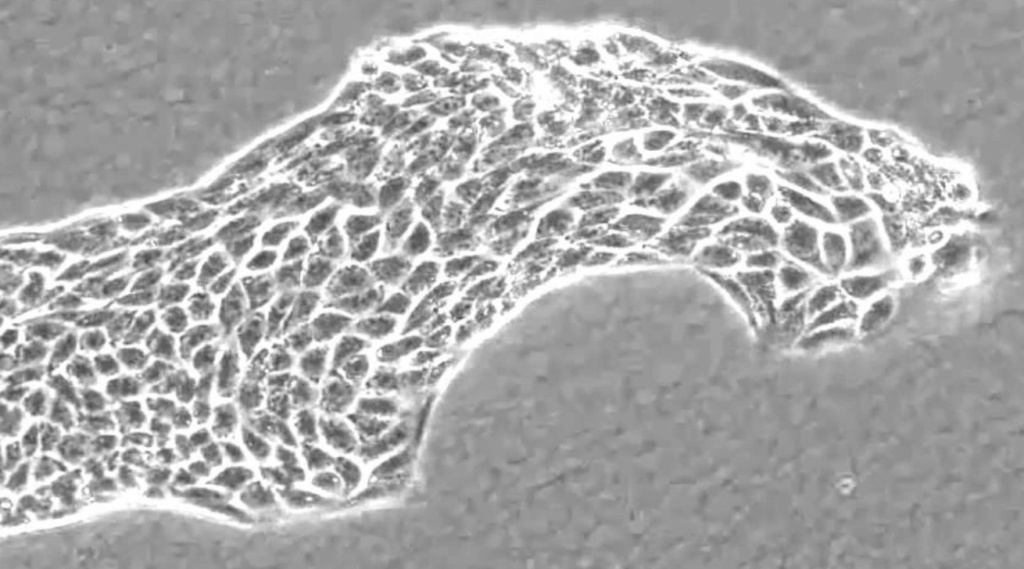
In recent years, evidence has accumulated confirming the importance of such intercellular communication and the molecular mechanisms involved. The addition of cells in an evolutionary trajectory from unicellular to multicellularity requires the production of precise new information of where the new cells have to be located in the evolving organism.
Processes such as tissue formation during embryonic development wound healing, and immune responses, all require the orchestrated movement of cells in particular directions to specific locations. Errors during this process have serious consequences, including intellectual disability, vascular disease, tumour formation and metastasis.
Eukaryotic cell migration typically is far more complex and can consist of combinations of different migration mechanisms. It generally involves drastic changes in cell shape which are driven by the cytoskeleton, for instance, a series of contractions and expansions due to cytoplasmic displacement.

When the tasks of labour are shared and/or divided in a multicellular organism, then the specific function to each cell must be purposefully defined, organized and assigned, in order to permit to build tissues, organs, organ systems, to organisms.
Epigenetics is a reference to a whole host of mechanisms that affect cellular function, tissue-specificity, and morphogenesis above and beyond the genetic code specified in the linear sequence of DNA nucleotides.
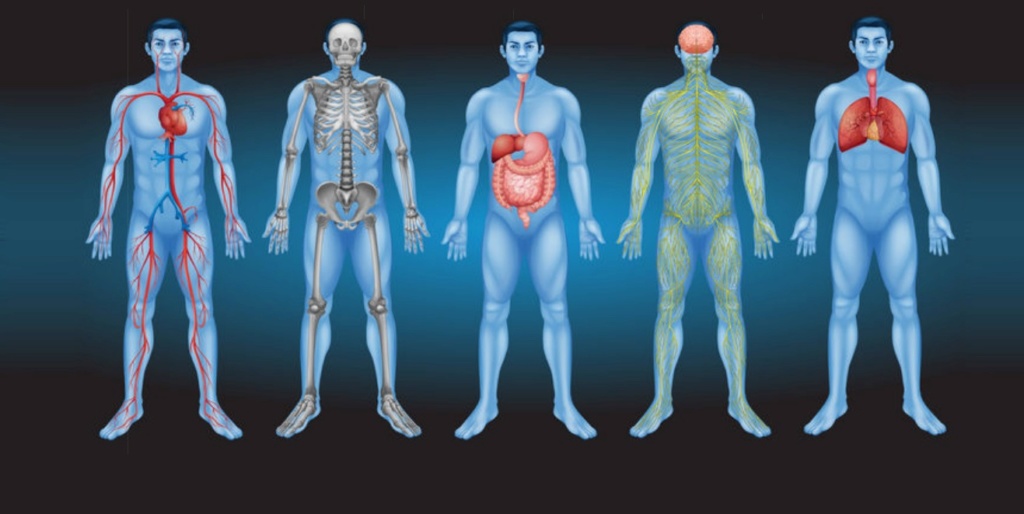
Each Cells needs to be in the right position and place in the body. This is crucial. Limbs like legs, fins, eyes etc. must all be placed at the right place.
It is awe inspiring to imagine, that the instructional information stored in one single fertilized egg can give rise to a human adult body, developing into an organism made of 37 trillion cells, perfectly organized, each cell at the right place, emerging amongst 200 different ones, organizing into tissues, organs etc.
Fingers are always at the tips of our hands, never in the middle; our eyes are always in our heads, not in our toes or gut. This creation of ordered form involves precisely coordinated cell growth, cell migration, and cell death.
This video is a powerful tool to demonstrate why micro does not lead to macroevolution, why Darwins Theory of evolution does not withstand scrutiny and the enormous biological
challenge that would have had to be overcome to go from unicellular to multicellular life.
https://www.youtube.com/watch?v=MdjjqBI_EzI
Has Paley's Watchmaker argument been debunked?
https://reasonandscience.catsboard.com/t2860-has-paley-s-watchmaker-argument-been-debunked
Some requirements for transition to multicellularity: 10.00
1, Cell differentiation: 13.32
2. Morphogenesis 18.15
3. Defining each Cell's specific function 25.40
4. Cell migration 27.07
5. Connecting adjacent cells 31.53
6. Cell-Cell communication 33.24
7. Adaptation & microevolution 43.35
8. The genetic program limiting body growth 47.12
9. Gene regulatory network 50.28
10. Setting up sexual reproduction 58.34
Endnotes: 1.02.54

William Paley was an English Christian apologist, and philosopher. He is best known for his natural theology exposition of the teleological argument for the existence of God which made use of the watchmaker analogy.
He believed that the complexity and perfection of nature were powerful arguments for the existence and attributes of God.

Paley famously made an argument by analogy of organisms as watches and of God as a watchmaker, according to which complex structures, like a watch, could not emerge accidentally but require the existence of a rational designer, in this case a watchmaker

Paleys watchmaker analogy is a classic: Paley writes: In crossing a heath, suppose I pitched my foot against a stone, and were asked how the stone came to be there, I might possibly answer, that, for any thing I knew to the contrary, it had lain there for ever: nor would it perhaps be very easy to shew the absurdity of this answer.

But suppose I had found a watch* upon the ground, and it should be enquired how the watch happened to be in that place, I should hardly think of the answer which I had before given, that, for any thing I knew, the watch might have always been there.

Yet why should not this answer serve for the watch, as well as for the stone? Why is it not as admissible in the second case, as in the first? For this reason, and for no other, that, when we come to inspect the watch, we perceive (what we could not discover in the stone) that its several parts are framed and put together for a purpose,
that they are so formed and adjusted as to produce motion, and that motion so regulated as to point out the hour of the day; that, if the several parts had been differently shaped from what they are, of a different size from what they are, or placed after any other manner, or in any other order, than that in which they are placed, either no motion at all would have been carried on in the machine, or none which would have answered the use, that is now served by it.

Without knowing about biology as we do today, Paley made observations, which are spot on, and have astounding significance and correctness, applied to the reality of molecular biochemistry. Let's list the points he mentioned again. If the parts are differently shaped, differently sized, placed after any other manner, or in any other order no motion would be the result.

the individual parts of the watch must be designed in order to be shaped and sized correctly, so they require an intelligent designer.

Each part must be complementary in order to be able to be placed at the right place and the assembly also requires the right sequence. The same is the case in a multicellular organism. This principle is called structural complementarity and is the very essence of biomolecular recognition.

In the same sense as in a watch, one part like an apoprotein without its substrate or prosthetic group must precisely fit and be able to interact with other parts. In biology, structural complementarity is the significant clue to understand the functional properties of biological systems.
Biological systems, from the macromolecular level to the cellular level, operate via specific molecular recognition mechanisms based on structural complementarity: A sperm, for example, recognizes an egg. All these interactions involve structural complementarity between molecules.

So according to the modern evolutionary claims, the watchmaker argument has been refuted by the theory of evolution.

Richard Dawkins borrowed the name from Paley for the title of his 1986 book The Blind Watchmaker, which attacked the idea that complex life needed a religious explanation. And still, today, when the watchmaker analogy is invoked, the immediate answer is: The analogy has been refuted through evolution by natural selection.
But has it? The emergence of multicellularity was ostensibly a major evolutionary leap. Indeed, most biologists consider it one of the most significant transitions in the evolutionary history of Earth’s inhabitants.
We will address this question by giving at what hurdles unicellular organisms would have had to overcome in order to evolve into multicellular organisms.

How do changes in development create new body architectures? Are heritable changes enough, given the constraints imposed by the necessity of the organism to survive as it develops? These questions are critical in molecular
biology, physiology, cell biology, genetics, anatomy, cancer research, neurobiology, immunology, ecology, and evolutionary biology.
The study of development has become essential for understanding all other areas of biology. In turn, the many advances of molecular biology have finally given elucidating answers. Have they been predicted correctly and in a satisfactory manner by Darwin's theory of evolution? The advance of science has it made possible to solve the riddle.
The black box has been opened. Has the evidence led to confirm evolutionary predictions, or have they have been falsified? Evolution would have had to go in a gradual slow, increasing manner from one eukaryotic cell to an organism with two cells, and so on, to get in the end an organism with millions, billions, or even trillions of precisely differentiated cells like the human body with 37 trillion cells.

Let's suppose there were unicellular organisms, eukaryotic so-called protozoans, and evolutionary pressure to go from one to two cells. What and how many mutations would be required in the genome and have to be selected and fixed? Mutations would have to provide the selection of a considerable number of new internal cell functions and created NEW information for these to emerge.

There would have to be set up a way to divide the tasks and assignments of cooperation to consume energy, survive, and reproduce. But how could just this apparently tiny, but in reality huge step in the molecular world have occurred? And why should it have? How did cells supposedly make the transition from life as single cells to split tasks, set up the information to start existing as bicellular organisms,

and associate and cooperate as teams such that they work as a single, cohesive unit? The change would englobe the modification of shape, divide functions and assign to each cell special tasks, structure, and entirely new features of development, ways to connect and stay connected to each other, communicate, and cooperate in order to start doing things in a combined manner.

And all this transition would have to be advantageous during each transitional step, resulting in an organism having an architecture where specialization of each cell in their functions would provide an advantage of survival. If they would duplicate and stick together, without specializing, they would just remain a group of cells or a colony.

Evolutionary literature claims that such transition supposedly occurred 25 times independently, which is called convergent evolution since cell-to-cell adhesion or communication is achieved in various ways in plants, animals, fungi, and algae.
1. Set up cell type and phenotype of the new cells
2. Set up morphology, size and shape of each new cell, and how its new agglomeration leads to new functional structures like tissues and organs organized in a non-random, functional manner.
3. Defining each cell's specific function after differentiation
4. Cell migration, set up and define the position and place in the body of each cell type.
5. Set up specific cell adhesion mechanisms for each cell type
6. Set up Cell-Cell communication channels and signalling networks
7. Set up mechanisms for adaptation and homeostasis specifically for the new emerging multicellular organism
8. The genetic program limiting body growth and cell cycle counter
9. Set up of the gene regulatory network orchestrating gene expression
10. Setting up sexual reproduction

As evidence reveals, a considerable number of innovations would have to be implemented, as listed above. The technical demands of multicellularity are extraordinary. Cells that commit to living together need a whole new set of tools. They have to come up with ways of sticking together, communicating, and sharing oxygen and food.
They also need a master developmental program, a way to direct specific cells to take on specialized jobs in different parts of the body. And a self-destruction program is necessary. The process is called apoptosis. The genes participating in adhesion in animals might have been extant in the unicellular ancestor of animals.
Most of the domains typically found in animals are present for example in choanoflagellates. However, the function of such a diverse set of adhesion molecules in a unicellular organism is not yet known. There are more demands and requirements which will not be elucidated in this video.

Why did cells not remain single-celled? In Astrobiology magazine, the answer to this question is usually cooperation, as cells benefitted more from working together than they would from living alone. Hummm. Really ??!! An article in Science magazine in June 2018 admitted that the transition to multicellular life as momentous as any in the history of life, until recently we had no idea how it happened.
The gulf between unicellular and multicellular life seems almost unbridgeable, so they confessed. If so, does the explanation above not seem, let's say, a little bit simplistic and naive? or, worse, even pseudo-scientific?

In the paper: The Origins of Multicellularity, the author writes: My line of reasoning is that the first step in the evolution of multicellularity was a size increase due to an accident, in other words, a mutation that prevents the daughter cells from separating. If the larger cell mass has any advantages, such as ensuring the safety of the germline by producing a protected internal environment, then natural selection will see to it that the novelty is retained.
Cells require ultrasophisticated proteins that keep them together. Arguing that an accident did hold them somehow together seems nothing more than just so guesswork. We will give now a closer look at the mentioned innovations.

Cell differentiation. That is already a huge venture. Stem cells can generate transit cells and, finally, differentiated cells. In most cases, stem cells can produce more than one type of differentiated progeny (multi- or totipotent). So the transition from a Cell containing just the information to make one cell, to a cell with two or more differentiated Cells would have to occur, sharing tasks.

Now let us suppose early earth was populated by unicellular life forms, like protozoans. Then the transformation to multicellular organisms began to evolve. Pluripotent cells would have had to become able to produce specialized cells, where each one of them would have had function only together with all other cells, like ten different types of cells forming blood.
This differentiation had to be organized and coordinated in advance on a systems level above the individual cell level. Foresight is required to know the end function of blood in a joint venture of all required individual blood cell types.

Blood is only functional if it carries all ten different cell types, which have each different, vital functions for life. Platelets, for example, are essential components of blood whose function is to stop bleeding by clumping and clotting blood vessel injuries.

T cells are essential for human immunity. They orchestrate an immune response and play important roles in all arms of immunity.

Red blood cells are the most common type of blood cells and essential for the vertebrate organism's delivering oxygen to the body tissues—via blood flow through the circulatory system.

At least six of the different cell types in the blood are life essential. If one is missing, life using cardiovascular systems is not possible. Had these blood types not to emerge together, to make the life of a multicellular organism like vertebrates possible?
Moreover, Vascular cells have also been shown to be part of the niche that controls the maintenance and differentiation of hematopoietic stem cells which form all blood cells and reside in the bone marrow. This forms an entire interdependent system where bones had to be existent, providing bone marrow, the place where blood and vascular formation takes place.

Most blood types require intermediate steps in their development until becoming final cell types used in blood. These intermediate cell types have no function in the organism. Why would evolution produce them or natural selection select them? We are talking about an enormous number of mutations required to produce each of the ten different blood cells.
Many cells play many roles during development, going through stages that are no longer seen in adulthood. How can natural selection account for the emergence of these cells? The role of some cell types is to activate specific genes in nearby cells, and once they accomplish their task, they die. This is clear evidence of an orchestrated, pre-programmed process.

I think Byron Bledsoe brings it pretty well to the point in this quote. Irreducible complexity is powerful evidence of creation, and the vascular system composed of veins and blood cells, and bone marrow where hematopoietic stem cells are produced, is an interdependent system, and so a very illustrative example. But how does science explain the origin of the blood vascular system?

It is evident that the authors in the rather recent paper cited above from 2013, Evolutionary origins of the blood vascular system and endothelium, evolution is assumed, but the authors have nothing to offer besides declarations of vague guesses and assertions based on evolutionary beliefism.

Morphogenesis

Morphogenesis depends on a serious of factors, amongst it defining the size and shape of each cell, which has to organize into functional structures. Differentiated cells are not randomly distributed. Rather, they are organized into intricate tissues and organs. During development, cells divide, migrate, and die; tissues fold and separate.
Genetic and epigenetic information has to be set up to define morphology, function, size and shape of each new cell, and how its new agglomeration leads to new functional structures like tissues and organs organized in a non-random, functional manner.

Cells come in all sorts of forms and sizes, and that is defined based on the functions they will exercise.

In an article in the magazine wired, HOW LIFE MADE THE LEAP FROM SINGLE CELLS TO MULTICELLULAR ANIMALS, the authors write: After the appearance of microbes, life got more complicated. Cells organized themselves into new three-dimensional structures. They began to divide up the labour of life so that some tissues were in charge of moving around, while others managed eating and digesting.
They developed new ways for cells to communicate and share resources. These complex multicellular creatures were the first animals, and they were a major success. Soon afterwards, roughly 540 million years ago, animal life erupted, diversifying into a kaleidoscope of forms in what’s known as the Cambrian explosion.

The longest-necked animals of all time were the extinct sauropod dinosaurs, some of which had necks 14 meters long. In these animals, the neurons that comprised the recurrent laryngeal nerve were at least 28 meters long. In the longest sauropods, these neurons may have been 40–50 meters long, probably the longest cells in the history of life.
They were sensory neurons that connected receptors in the skin of the extremities with interneurons in the brainstem, a pattern of neural architecture that is present in all extant vertebrates.

The biggest single-celled organism can be up to 20cm across – Syringammina fragilissima’s one cell branches out into a network of tubes extending over ten centimetres ( or 3.9 inches). As it grows, the deep sea-dwelling creature oozes slime onto the sediment, solidifying its structure.

The size and form of a hospital are totally different than of a residential house. What defines its structure, is the scaffold structure, which has to be defined precisely taking the specific demands in consideration.

Shape and form of cells have to be defined in accordance with the function that each cell type will perform. A neuron requires a totally different form and shape than a red blood cell.

In cells, actin, actomyosin and microtubules that form the cytoskeleton exercise the same function as the scaffold structure of a house or a large building, defining shape and form.
For cells to function properly, they must organize themselves and interact mechanically with each other and with their environment. They have to be correctly shaped, physically robust, and properly structured internally.

Many have to change their shape and move from place to place. All cells have to be able to rearrange their internal components as they grow, divide, and adapt to changing circumstances. These spatial and mechanical functions depend on this remarkable system of filaments.

The cytoskeleton’s varied functions depend on the behaviour of three families of protein filaments—actin filaments, microtubules, and intermediate filaments. Microtubules are very important in a number of cellular processes. They are involved in maintaining the structure of the cell and provide a platform for intracellular macromolecular assemblies through dynein and kinesin motors.

They are also involved in chromosome separation (mitosis and meiosis) and are the major constituents of mitotic spindles, which are used to pull apart eukaryotic chromosomes. The dynamic cytoskeletal and membrane system that governs morphological patterning in cells is fundamental.
This system enables eukaryotes to package signalling molecules in endosomes, tiny vacuoles that bud from surface membranes, and transport them through the cell by means of molecular motors at rates much faster than would be possible by diffusion alone
As a result, eukaryotic cells can change shape or physiology in response to molecular signals, permitting processes such as locomotion of amoebas, and permanent cell differentiation. The dynamic cytoskeleton and membrane system of eukaryotic cells open up possibilities of size, structure, function, and development not available to prokaryotic organisms.

Mitotic cell division is the most fundamental task of all living cells. Cells have the intricate and tightly regulated machinery to ensure that mitosis occurs with appropriate frequency and high fidelity. If someone wants to explain the origin of eukaryotic cells, the arise of mitosis and its mechanism and involved cell organelles and proteins must be elucidated.
The centrosome plays a crucial role: it functions as the major microtubule-organizing center and plays a vital role in guiding chromosome segregation during mitosis. These architecturally perfect structures are essential in many animal cells and plants.

Defining each Cell's specific function Genes define the amino acid sequence that makes proteins, and specific sets of proteins define tissue-type. There are no proteins for hemoglobin in neuronal cells and there aren't proteins that make voltage-gated ion channels in red blood cells.
These functional elements of the cells, along with other cell-specific proteins, are responsible for making a neuron a neuron and a red blood cell a red blood cell. Yet, both cell-types have the genes for each of these cell-specific proteins, in fact every eukaryotic cell with a nucleus in a multicellular organism contains a full set of all tissue-specific genes as part of the full genome.
Tissue-specificity must then arise from how genes are differentially expressed, meaning that in some cell lineages certain sets of genes are silenced, while in others they are expressed – leading one line of progenitor stemcells to differentiate into neurons and another to differentiate into erythrocytes, or any of the other approximately 200 different cell types of the human body.

Another issue that has to be explained is individual, and a more advanced stage, collective cell migration which has a key role during morphogenesis and during wound healing and tissue renewal in the adult. It is a central process in the development and maintenance of multicellular organisms.
In addition to displaying a coordinated migratory behaviour, collectively migrating cells move more efficiently than if they migrated separately, which indicates that a cellular interplay occurs during collective cell migration.

In recent years, evidence has accumulated confirming the importance of such intercellular communication and the molecular mechanisms involved. The addition of cells in an evolutionary trajectory from unicellular to multicellularity requires the production of precise new information of where the new cells have to be located in the evolving organism.
Processes such as tissue formation during embryonic development wound healing, and immune responses, all require the orchestrated movement of cells in particular directions to specific locations. Errors during this process have serious consequences, including intellectual disability, vascular disease, tumour formation and metastasis.
Eukaryotic cell migration typically is far more complex and can consist of combinations of different migration mechanisms. It generally involves drastic changes in cell shape which are driven by the cytoskeleton, for instance, a series of contractions and expansions due to cytoplasmic displacement.

When the tasks of labour are shared and/or divided in a multicellular organism, then the specific function to each cell must be purposefully defined, organized and assigned, in order to permit to build tissues, organs, organ systems, to organisms.
Epigenetics is a reference to a whole host of mechanisms that affect cellular function, tissue-specificity, and morphogenesis above and beyond the genetic code specified in the linear sequence of DNA nucleotides.

Each Cells needs to be in the right position and place in the body. This is crucial. Limbs like legs, fins, eyes etc. must all be placed at the right place.
It is awe inspiring to imagine, that the instructional information stored in one single fertilized egg can give rise to a human adult body, developing into an organism made of 37 trillion cells, perfectly organized, each cell at the right place, emerging amongst 200 different ones, organizing into tissues, organs etc.
Fingers are always at the tips of our hands, never in the middle; our eyes are always in our heads, not in our toes or gut. This creation of ordered form involves precisely coordinated cell growth, cell migration, and cell death.
Last edited by Admin on Sun May 26, 2019 10:13 pm; edited 53 times in total






































