


ONTOGENY AND PHYLOGENY OF BLOOD CELL PROGENITORS (HEMOCYTOBLASTS) 3
Blood cells are produced continuously in the mature animal. More than most other cell types, their rate of formation fluctuates. Responsible for many metabolic functions and, in particular, the immune response, blood cell formation has to be upregulated on demand upon injuries and pathogen invasion.Although mitotic division of mature hemocytes (e.g., plasmatocytes) has been observed in most animal taxa, the majority of blood cells appear to derive from self-renewing populations of multipotent stem cells [termed hemocytoblast, or hemocyte stem cell (HSC), in vertebrates and hemocyte progenitors in invertebrates] that are housed in specialized hematopietic organs. This section provides a brief comparative overview of the structure of these hematopoietic organs. Subsequently, developmental and molecular aspects of early hematopoiesis (that is, the formation of hemocytoblasts) are discussed for vertebrates and Drosophila, the one invertebrate for which recent studies have shed light on blood development.
Structure of Hematopoietic Organs
Hematopoietic organs have been described for all major taxa of coelomate animals. In invertebrates, they are typically mesenchymal or gland-like structures 4 attached to the lining of blood vessels and/or the coelomic cavity. 5 , specialized domains within the mesothelium 6 show higher rates of proliferation and bud off hemocytes into the lumen of the coelom or blood vessels (Figure 3)
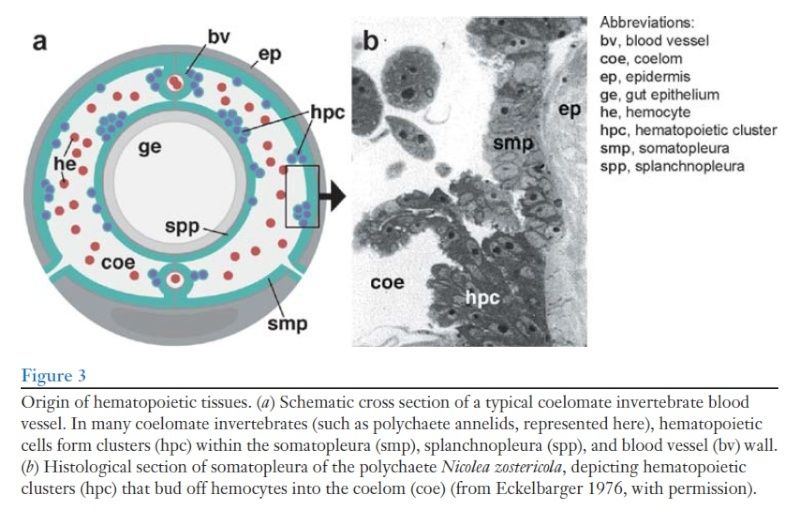
The same origin of hemocytes from mesothelial cells lining the coelom/blood vessels has been observed in other invertebrates, including lower deuterostomes 7. This hematopoietic mechanism may give us a glimpse into the origin of the close ontogenetic relationship between hemocytes and vascular cells. Thus, for Drosophila a common progenitor (prior to its last round of division) gives rise to both vascular and hemocyte progenitors , and the same seems highly likely in vertebrates as well. In vertebrates, suspected common progenitors of endothelial and blood progenitors were termed hemangioblasts . It is reasonable to assume that phylogenetically, hemangioblasts originated as bi- or pluripotent cells populating the coelomic epithelium of a primitive ancestral invertebrate. More detailed studies of the origin of hemocytes in polychaetes and other simple coelomates, which may have retained primitive aspects of the bilaterian ancestor, may be highly informative in regard to the evolutionary origin of hematopoietic organs.
Why is it reasonable to assume that ? This are unwarranted assumptions, and the author does not inform why that should be a reasonable assumption. Furthermore, this key sentence acknowledges that science has no clue about the evolutionary origin of pluripotent cells.
We encounter a more complex hematopoietic mechanism in oligochaetes, some molluscs, arthropods, and ascidians. In these taxa, hematopoietic stem cells moved out of the mesothelium and coalesced in compact hematopoietic organs called lymphoid organs or lymph glands (Figure 4). Lymph glands are typically attached to the coelomic wall or large blood vessels. In oligochaetes and insects, lymph glands form a metameric pattern of encapsulated organs flanking the wall of the dorsal blood vessel (Figure 4b). These invertebrate lymph glands consist of spongy mesenchymal masses of cells, 8 many of which are mitotically active and give birth to various types of hemocytes that initially fill the lacunae within the center of the gland before moving out into the lumen of the coelom or hemolymph space (Figure 4c).
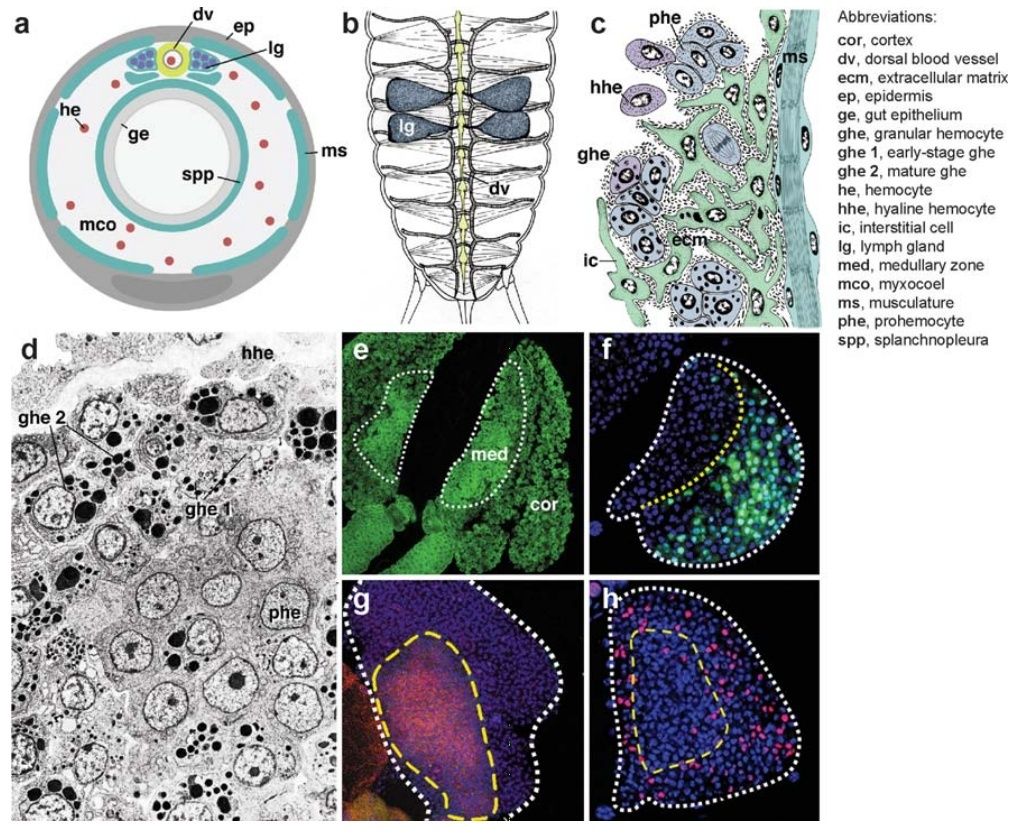
Figure 4 Hematopoietic organs (lymph glands) in invertebrates.
(a) Schematic cross section of arthropod showing a close association of lymph gland (lg) and dorsal blood vessel (dv).
(b) Schematic dorsal view of insect, showing dorsal blood vessel (dv) and attached bilateral, segmentally arranged lymph glands (lg) (after Hoffmann et al. 1979). The location of lymph glands along the anteroposterior axis varies among insects: In cricket, shown here, lymph glands occur in the abdomen; in Drosophila, they are found around the anterior portion of the dorsal vessel in the thorax.
(c) Line drawing of section of insect lymph gland (after Hoffmann et al. 1979). Attached to dorsal muscles (ms) and dorsal vessel (not shown), the lymph gland consists of immature prohemocytes (phe) that give rise to different lineages of hemocytes (hhe, hyaline hemocytes; ghe, granular hemocytes). Mesodermal interstitial cells (ic) may represent stroma or the early stage of blood progenitors.
(d ) Electron micrograph of hematopoietic organ of ascidian (from Ermak 1976, with permission). Note the layered organization, with prohemocytes (phe) at the lower right, early stages of granular hemocytes (ghe 1) in the center, and more mature granular cells (ghe 2) at the upper left.
(e–h) Confocal images of Drosophila larval lymph gland (from Jung et al. 2005, with permission). (e) Global nuclear staining, depicting tightly clustered immature prohemocytes in the center (med, medullary zone), surrounded by more loosely packed differentiating hemocytes (cor, cortex).
( f ) Hemocyte differentiation marker (P1; green) labels exclusively cortex of lymph gland.
( g ) Adhesion molecule DE-cadherin (red ) is expressed at high levels in the medullary zone.
(h) Proliferation (BrdU; red ) is more pronounced in the cortex.
Similar structures have also been described for mollusks [e.g., the amebocyte-producing organ in gastropods, the “white glands” of cephalopods located close to the eye and crustaceans (the lymph glands along the vessels near the eyes and proximal appendages; reviewed in Bauchau 1981). In ascidians, lymph glands (described as blood-forming nodules) are grouped around the transverse bars of the pharyngeal basket, generally assumed to represent the phylogenetic forebear of the gill apparatus of vertebrates . Invertebrate lymph glands contain mostly proliferating prohemocytes but also contain differentiated blood cells such as plasmatocytes. A zonation is often visible (Figure 4d– h), such that the undifferentiated hemocyte progenitors clump together, and more differentiated hemocytes form an outer layer around them. A prominent stroma, akin to the network of fibroblasts and capillaries found in the hematopietic tissue of vertebrates, is missing in invertebrates. In some instances, scattered muscle cells or undifferentiated mesenchymal cells penetrate the lymph glands; a basement typically covers the gland at its outer surface . The bone marrow is the hematopoietic organ in all vertebrates but fishes, in which hematopoiesis occurs in the kidney. Osteoblasts (bone-forming cells) form a layer, termed endosteum, at the interface between the mineralized bone and the bone marrow contained within its center. Blood vessels, capillaries, and wide, endothelium-bound spaces called venous sinuses branch throughout the bone marrow (Figure 5a–c).
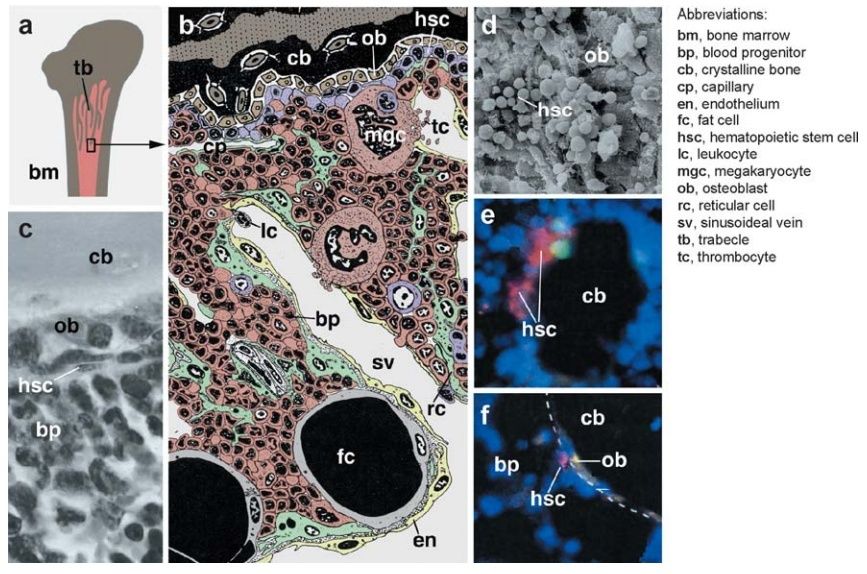
Figure 5 Hematopoiesis in vertebrate bone marrow.
(a) Marrow (bm) lies in the center of long bones. Branched processes of bone, called trabecles (tb), protrude into the marrow and greatly enlarge the surface at which bone and marrow contact each other.
(b) Colored line drawing of bone marrow section. Trabecle formed by crystalline bone (cb) is seen at the top of the picture. Bone-forming cells (ob, osteoblasts) are located within the bone matrix but also form an epithelial layer (endosteum) at inner bone surface. The center of the bone marrow is formed by endothelially (en) lined capillaries (cp) and sinusoideal veins (sv) that are connected with each other and the bone surface by a meshwork of reticular cells (rc; green). Reticular cells form the stroma of the bone marrow, which contains proliferating and differentiating clusters of hematopoietic stem cells (HSCs) (denoted in figure by hsc; violet) and blood progenitors (bp; brown) as well as postmitotic maturing blood cells. HSCs are enriched in the zone that contacts osteoblasts; this zone is believed to constitute the HSC niche. Differentiating blood cells are seen to enter the circulation; these include thrombocytes (tc), produced by the fragmentation of large megakaryocytes (mgc), and leukocytes (lc).
(c) Histological section of human bone marrow showing the interface of bone matrix (cb), covered by osteoblasts (ob), with HSCs and blood progenitors (bp) (from Balduino et al. 2005, with permission).
(d ) Scanning electron micrograph of inner bone surface with osteoblasts (ob) and HSCs (from Balduino et al. 2005, with permission).
(e, f ) Confocal section of embryonic mouse bone marrow. Trabecle of bone (cb) is surrounded by osteoblasts (ob) and bone marrow (blue; global nuclear labeling). Panels e and f from Zhang et al. 2003, with permission. (e) HSCs are labeled in pink by stem cell marker Sca; the green label represents BrdU-positive proliferating cells.
( f ) Contact of osteoblasts (ob), marked by expression of N-cadherin ( yellow), with HSCs (pink; labeled with BrdU).
Endothelial cells, osteoblasts, and stromal (also called reticular) cells that crisscross the space between vessels and endosteum form a threedimensional scaffold that houses clusters of blood-forming cells. This scaffold provides a complex microenvironment that, by means of cell-membrane-bound and secreted factors, controls the determination and proliferation of the different blood cell lineages. The multipotent hematopoietic stem cells (HSCs) that seed the bone marrow in the late embryo settle at the outer (subendosteal) layer in contact with the osteoblasts (Figure 5d–f ); these latter cells, forming the stem cell niche of the bone marrow, emit signals that maintain HSCs in their noncommitted stem cell mode. HSC-derived cells that lose contact with the osteoblast layer progress toward the next stage, that of a committed progenitor for lymphoid cells, red blood cells, thrombocytes, (neutrophile) granulocyte/monocytes, basophile granulocytes, or eosinophile granulocytes. These different progenitors are then found nearer the center of the bone marrow, where they proliferate and form growing colonies of maturing blood cells (Figure 5b,c). Once matured, blood cells become capable of crossing the endothelium into the bloodstream. Lymphoid progenitors leave the bone marrow at an immature state and populate the thymus and lymphoid organs.
Hematopoiesis and Hematopoietic Organs in Arthropods 1
Most animals possess populations of cells that are not fixed in place as part of an organ, but that roam within the body cavities to fulfill functions involved with the immune response, cell degradation and replacement, and the distribution of nutrients and gases. The blood cells, or hemocytes, of the vertebrate body, which comprise many different classes each responsible for one of the functions mentioned above, represent the best known example. All invertebrates which possess a body cavity and, to various degrees, a vascular system also have hemocytes of different types. Even in many animals without a body cavity, including cnidaria, sponges and flatworms, one observes motile cells that primarily function as pools of pluripotent stem cells. Such cells may represent the ancestral type of motile cell from which the diverse, specialized classes of hemocytes of higher animals evolved (van de Vyver, 1981).
Hemocytes typically have a lifespan of days to weeks, and need to be replaced constantly. Furthermore, given that the demand for hemocytes (in particular those involved in the immune response, and the related processes of wound healing and regeneration) varies over time, the rate at which hemocytes are produced must be able to adapt to these variable conditions. Blood cells are produced in specialized tissues, called hematopoietic organs. Hematopoietic organs generate blood cells in high numbers throughout the life time of the animal, implying that hematopoietic organs must contain, or be supplied with, self renewing stem cells. The study of hematopoiesis thus addresses a central issue of stem cell biology, namely the “decision” of an undifferentiated cell to either continue to proliferate, or to leave this state to initiate differentiation. We will in the following start out by briefly summarizing pertinent facts about blood cell development among the invertebrates and then provide more details illustrating the architecture and proliferative function of hematopoietic organs in the arthropods.

Origin of hematopoietic tissues.
A: In many coelomate invertebrates (such as polychaete annelids represented here as schematic cross section) hemocytes are produced in hematopoietic centers within the somatopleura, splanchnopleura, and blood vessel wall
B: Histological section of somatopleura of the polychaete Nicolea zostericola, showing hematopoietic center from where groups of hemocytes bud off into the coelom (Scale bar: 18 m; from Eckelbarger, 1976, with permission).
C: Schematic cross section of insect showing hematopoietic lymph gland (lg) closely associated with the dorsal blood vessel.
D: Histological section of the compact hematopoietic organ (lymph gland) of lepidopteran Manduca sexta (Scale bar: 50 m; from Nardi et al., 2003, with permission).
In these cases, progenitors of hemocytes and endothelial/mesothelial cells form one and the same cell type. In other cases, hematopoietic cells form mesenchymal clusters (“nodules”) or gland-like structures (“lymph glands”) that are separate from the vascular linings, but always remain in close contact with them (Fig.1C, D). In Drosophila, where the development of an invertebrate hematopoietic organ was studied, it became clear that a common progenitor (hemangioblast) gives rise to both vascular cells and hemocytes (Mandal et al., 2004), which is similar to the situation in vertebrate embryos (Choi et al., 1998; Teixera et al., 2011). In the Drosophila embryo, vascular progenitors undergo a mesenchymal-epithelial transition to form the lining of the blood vessels; blood progenitors assemble into mesenchymal clusters that remain attached to the basal surface of the vascular epithelia. These clusters proliferate rapidly and develop into the lymph glands of the larva. Lymph glands have been observed in many insects. They form metamerically reiterated encapsulated structures that flank the wall of the dorsal blood vessel (Cuenot, 1897; Hoffmann et al., 1979, 1979). Similar structures have also been described for mollusks (e.g., the amebocyte producing organ in gastropods; Jeong et al., 1983; the “white glands” of cephalopods; Cowden and Curtis, 1981) and crustaceans (lymph glands along the vessels and over the foregut; reviewed inBauchau, 1981). In lower deuterostomes (e.g., ascidians), “blood forming nodules” are grouped around the transverse bars of the pharyngeal basket (Wright, 1981).
Hemocytes have been studied in a number of insects, crustaceans and chelicerates, with much controversy associated with the number of different hemocyte cell types observed and the terminology used to designate them. Thus, often researchers used different terms for the same type of cell; or, differences in cell structure that lead to different names merely reflected artefacts of fixation, or different stages during the development of the same cell. Attempts to simplify and unify the terminology have been made repeatedly. The more recent literature recognizes the following main cell types (reviewed in Jones, 1970; Rowley and Ratcliffe, 1981; Pelc, 1986; Lavine and Strand 2002; Ribeiro and Brehelin, 2006): prohemocytes, plasmatocytes, granulocytes, oenocytoids, and spherule cells (also called adipohemocytes). (1) Prohemocytes (Fig.2D) are small, rounded cells with a high nucleus:cytoplasm ratio and no specialized cytoplasmic organelles; these cells represent the immature hemocyte progenitors, or stem cells. All other blood cell types are derived from prohemocytes. (2) Plasmatocytes (Fig.2A) have more cytoplasm than prohemocytes, and their membrane forms ruffles and filopodia. Plasmatocytes are phagocytotically active, ingesting cellular debris during normal development, and engulfing microorganisms during the immune response. Plasmatocytes are also able to differentiate into large, flattened cells that arrange themselves around foreign bodies that have gained access to the body cavity, forming a capsule around these invaders (Fig.2J). These enlarged, flattened plasmatocytes were called lamellocytes in Drosophila and other insects (Rowley and Ratcliffe, 1981). (3) Granulocytes (Fig.2B) have electron-dense, regularly sized, acidophilic granula. Like plasmatocytes, granulocytes also act as phagocytes. Some insects, including Drosophila, lack granulocytes; here, plasmatocytes take on the function of granulocytes. The loss of granulocytes is apparently not evolutionary ancient; other Dipterans, such as Calliphora, possess granulocytes (Kaaya and Ratcliffe, 1982). (4) Oenocytoids (Fig.2E) are large, oval cells whose cytoplasm contains agglomerates of crystalline material and/or microtubules. The so called crystal cells of Drosophila (Fig.2C) correspond to the oenocytoids defined for other arthropods (Brehelin, 1982). Oenocytoids store prophenol oxidase, an enzyme that plays an important role in the innate immune response (see below). (5) Spherulocytes (Fig.2F) or adipohemocytes are blood cells with basophilic, variably sized and shaped granules. These cells may represent late stages in the differentiative pathway of phagocytic plasmatocytes. The range of blood cells characterized above is simpler, or modified, in some cases. For example, in Drosophila, we do not encounter granulocytes or spherulocytes. Crustaceans and many chelicerates lack oenocytoids; prophenol oxidase is produced by granulocytes (Zhang et al., 2006; Lin and Soderhall, 2011).
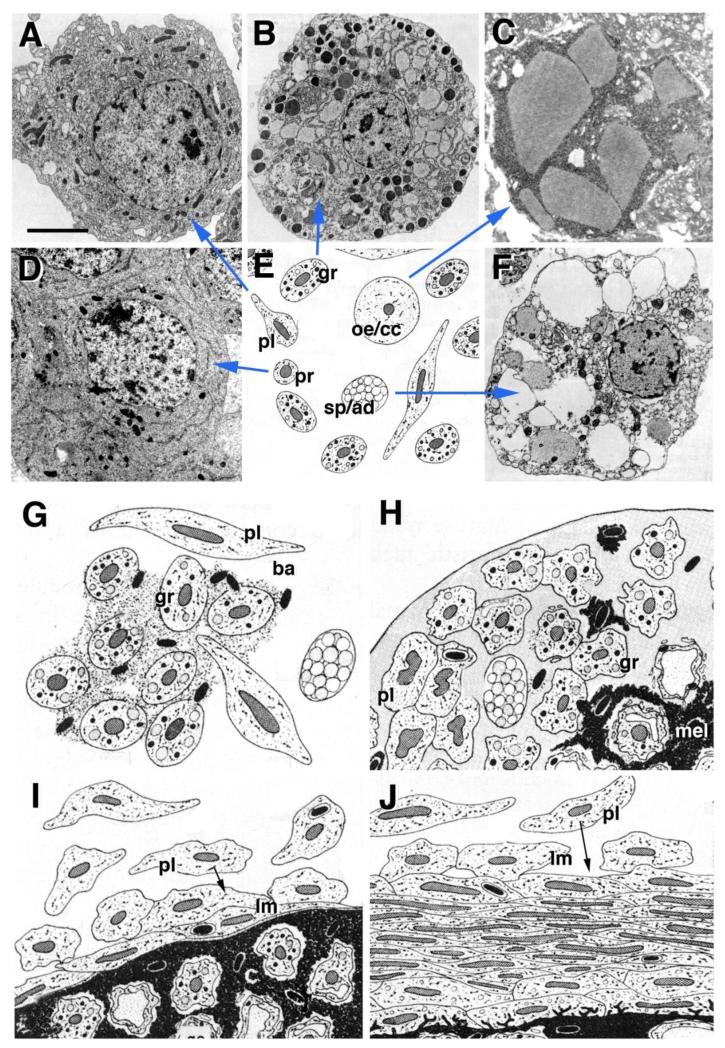
A-F: Types of arthropod hemocytes (summarized in E), including
(A) plasmatocytes (pl)(from Rowley and Ratcliffe, 1981, with permission),
(B) granulocytes (gr) (from Rowley and Ratcliffe, 1981, with permission),
(C) oenocytoids/crystal cells (oe/cr)(from Rizki and Rizki, 1984, with permission),
(D) prohemocytes (pr)(from Hoffmann et al., 1979, with permission), and
(F) spherulocytes/adipohemocytes (sp/ad) (from Rowley and Ratcliffe, 1981, with permission).
G-J: Schematic depiction of innate immune reaction carried out by different classes of hemocytes in response to invading microorganisms (ba bacteria; from Rowley and Ratcliffe, 1981, with permission). For detail, see text. Scale bar (A-D, F): 2μm.
Arthropod hemocytes mainly function during the innate immune response and wound healing (Rowley and Ratcliffe, 1981; Nappi et al., 2009; Jiang et al., 2010; Krzemien et al., 2010; Lin and Soderhall, 2011). Invading microorganisms attract plasmatocytes and granulocytes, which release reagents that entrap the invaders (Fig.2G, H). Products secreted by granulocytes also play a role in hemolymph coagulation. Plasmatocytes, differentiating as lamellocytes, form a capsule around the invaders (Fig.2I, J). Oenocytes release prophenol oxidase, which is converted into activated phenol oxidase by a cascade of enzymes that form part of the hemolymph, and that are activated by foreign molecules (e.g., microbial lipopolysaccharides). Phenoloxidase catalyzes a reaction by which melanin is synthesized (Fig.2H-J), and toxic metabolites (e.g., quinones) are released. Encapsulation and melanization act to neutralize the invading microbes or parasites.
How is bug blood different from our own? 2
The major difference between insect blood and the blood of vertebrates, including humans, is that vertebrate blood contains red blood cells. Insects and other invertebrates, on the other hand, have what is called hemolympha heterogeneous fluid that courses through their bodies, bathing all the internal tissues. Hemolymph is mostly water, but it also contains ions, carbohydrates, lipids, glycerol, amino acids, hormones, some cells and pigments. The pigments, however, are usually rather bland, and thus insect blood is clear or tinged with yellow or green. (The red color that you see upon squashing a housefly or fruit fly is actually pigment from the animal's eyes.)
Unlike the closed circulatory system found in vertebrates, insects have an open system lacking arteries and veins. The hemolymph thus flows freely throughout their bodies, lubricating tissues and transporting nutrients and wastes. Whereas the vertebrate circulatory system serves primarily to carry oxygen throughout the body, insects respire an entirely different waynamely, through tracheal tubes. In the case of the fruit fly Drosophila, for example, a series of tiny openings called spiracles line the impermeable outer skin of the fly, and these convey air directly to tracheal tubes that, in turn, convey air to the internal tissues.
Insects do have hearts that pump the hemolymph throughout their circulatory systems. Though these hearts are quite different from vertebrate hearts, some of the genes that direct heart development in the two groups are in fact very similar. The development and evolution of the vertebrate heart is currently the subject of much research.
1) http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3873168/
2) http://www.scientificamerican.com/article/how-is-bug-blood-differen/
3) http://materiais.dbio.uevora.pt/BD/Diferenciacao/Hemacytoblast.pdf
4) ( Mesenchymal stem cells, or MSCs, are multipotent stromal cells that can differentiate into a variety of cell types )
5) ( Refers to the main body cavity in most multicellular animals .A body cavity is any fluid-filled space in a multicellular organism other than those of vessels (such as blood vessels and lymph vessels). The term usually refers to the space located between an animal’s outer covering (epidermis), and the outer lining of the gut cavity, where internal organs developIn the simplest scenario (e.g., some polychaetes)
6) The mesothelium is a membrane composed of simple squamous cells that forms the lining of several body cavities: the pleura (thoracic cavity), peritoneum (abdominal cavity including the mesentery), mediastinum andpericardium (heart sac). Mesothelial tissue also surrounds the male internal reproductive organs (the tunica vaginalis testis) and covers the internal reproductive organs of women (the tunica serosa uteri). Mesothelium that covers the internal organs is called visceral mesothelium, while the layer that covers the body walls is called the parietal mesothelium. Mesothelium is the epithelial component of serosa.
7) Deuterostomes (taxonomic term: Deuterostomia; from the Greek: "mouth second") are any members of a superphylum of animals. Deuterostomia is a subtaxon of the Bilateria branch of the subkingdom Eumetazoa, within Animalia, and are distinguished from protostomes by their embryonic development; in deuterostomes, the first opening (the blastopore) becomes the anus, while in protostomes, it becomes the mouth. Deuterostomes are also known as enterocoelomates because their coelom develops through enterocoely.
8 Mesenchymal stem cells, or MSCs, are multipotent stromal cells that can differentiate into a variety of cell types, including: osteoblasts (bone cells), chondrocytes (cartilage cells), myocytes (muscle cells) andadipocytes (fat cells). This phenomenon has been documented in specific cells and tissues in living animals and their counterparts growing in tissue culture.
Blood cells are produced continuously in the mature animal. More than most other cell types, their rate of formation fluctuates. Responsible for many metabolic functions and, in particular, the immune response, blood cell formation has to be upregulated on demand upon injuries and pathogen invasion.Although mitotic division of mature hemocytes (e.g., plasmatocytes) has been observed in most animal taxa, the majority of blood cells appear to derive from self-renewing populations of multipotent stem cells [termed hemocytoblast, or hemocyte stem cell (HSC), in vertebrates and hemocyte progenitors in invertebrates] that are housed in specialized hematopietic organs. This section provides a brief comparative overview of the structure of these hematopoietic organs. Subsequently, developmental and molecular aspects of early hematopoiesis (that is, the formation of hemocytoblasts) are discussed for vertebrates and Drosophila, the one invertebrate for which recent studies have shed light on blood development.
Structure of Hematopoietic Organs
Hematopoietic organs have been described for all major taxa of coelomate animals. In invertebrates, they are typically mesenchymal or gland-like structures 4 attached to the lining of blood vessels and/or the coelomic cavity. 5 , specialized domains within the mesothelium 6 show higher rates of proliferation and bud off hemocytes into the lumen of the coelom or blood vessels (Figure 3)
The same origin of hemocytes from mesothelial cells lining the coelom/blood vessels has been observed in other invertebrates, including lower deuterostomes 7. This hematopoietic mechanism may give us a glimpse into the origin of the close ontogenetic relationship between hemocytes and vascular cells. Thus, for Drosophila a common progenitor (prior to its last round of division) gives rise to both vascular and hemocyte progenitors , and the same seems highly likely in vertebrates as well. In vertebrates, suspected common progenitors of endothelial and blood progenitors were termed hemangioblasts . It is reasonable to assume that phylogenetically, hemangioblasts originated as bi- or pluripotent cells populating the coelomic epithelium of a primitive ancestral invertebrate. More detailed studies of the origin of hemocytes in polychaetes and other simple coelomates, which may have retained primitive aspects of the bilaterian ancestor, may be highly informative in regard to the evolutionary origin of hematopoietic organs.
Why is it reasonable to assume that ? This are unwarranted assumptions, and the author does not inform why that should be a reasonable assumption. Furthermore, this key sentence acknowledges that science has no clue about the evolutionary origin of pluripotent cells.
We encounter a more complex hematopoietic mechanism in oligochaetes, some molluscs, arthropods, and ascidians. In these taxa, hematopoietic stem cells moved out of the mesothelium and coalesced in compact hematopoietic organs called lymphoid organs or lymph glands (Figure 4). Lymph glands are typically attached to the coelomic wall or large blood vessels. In oligochaetes and insects, lymph glands form a metameric pattern of encapsulated organs flanking the wall of the dorsal blood vessel (Figure 4b). These invertebrate lymph glands consist of spongy mesenchymal masses of cells, 8 many of which are mitotically active and give birth to various types of hemocytes that initially fill the lacunae within the center of the gland before moving out into the lumen of the coelom or hemolymph space (Figure 4c).
Figure 4 Hematopoietic organs (lymph glands) in invertebrates.
(a) Schematic cross section of arthropod showing a close association of lymph gland (lg) and dorsal blood vessel (dv).
(b) Schematic dorsal view of insect, showing dorsal blood vessel (dv) and attached bilateral, segmentally arranged lymph glands (lg) (after Hoffmann et al. 1979). The location of lymph glands along the anteroposterior axis varies among insects: In cricket, shown here, lymph glands occur in the abdomen; in Drosophila, they are found around the anterior portion of the dorsal vessel in the thorax.
(c) Line drawing of section of insect lymph gland (after Hoffmann et al. 1979). Attached to dorsal muscles (ms) and dorsal vessel (not shown), the lymph gland consists of immature prohemocytes (phe) that give rise to different lineages of hemocytes (hhe, hyaline hemocytes; ghe, granular hemocytes). Mesodermal interstitial cells (ic) may represent stroma or the early stage of blood progenitors.
(d ) Electron micrograph of hematopoietic organ of ascidian (from Ermak 1976, with permission). Note the layered organization, with prohemocytes (phe) at the lower right, early stages of granular hemocytes (ghe 1) in the center, and more mature granular cells (ghe 2) at the upper left.
(e–h) Confocal images of Drosophila larval lymph gland (from Jung et al. 2005, with permission). (e) Global nuclear staining, depicting tightly clustered immature prohemocytes in the center (med, medullary zone), surrounded by more loosely packed differentiating hemocytes (cor, cortex).
( f ) Hemocyte differentiation marker (P1; green) labels exclusively cortex of lymph gland.
( g ) Adhesion molecule DE-cadherin (red ) is expressed at high levels in the medullary zone.
(h) Proliferation (BrdU; red ) is more pronounced in the cortex.
Similar structures have also been described for mollusks [e.g., the amebocyte-producing organ in gastropods, the “white glands” of cephalopods located close to the eye and crustaceans (the lymph glands along the vessels near the eyes and proximal appendages; reviewed in Bauchau 1981). In ascidians, lymph glands (described as blood-forming nodules) are grouped around the transverse bars of the pharyngeal basket, generally assumed to represent the phylogenetic forebear of the gill apparatus of vertebrates . Invertebrate lymph glands contain mostly proliferating prohemocytes but also contain differentiated blood cells such as plasmatocytes. A zonation is often visible (Figure 4d– h), such that the undifferentiated hemocyte progenitors clump together, and more differentiated hemocytes form an outer layer around them. A prominent stroma, akin to the network of fibroblasts and capillaries found in the hematopietic tissue of vertebrates, is missing in invertebrates. In some instances, scattered muscle cells or undifferentiated mesenchymal cells penetrate the lymph glands; a basement typically covers the gland at its outer surface . The bone marrow is the hematopoietic organ in all vertebrates but fishes, in which hematopoiesis occurs in the kidney. Osteoblasts (bone-forming cells) form a layer, termed endosteum, at the interface between the mineralized bone and the bone marrow contained within its center. Blood vessels, capillaries, and wide, endothelium-bound spaces called venous sinuses branch throughout the bone marrow (Figure 5a–c).
Figure 5 Hematopoiesis in vertebrate bone marrow.
(a) Marrow (bm) lies in the center of long bones. Branched processes of bone, called trabecles (tb), protrude into the marrow and greatly enlarge the surface at which bone and marrow contact each other.
(b) Colored line drawing of bone marrow section. Trabecle formed by crystalline bone (cb) is seen at the top of the picture. Bone-forming cells (ob, osteoblasts) are located within the bone matrix but also form an epithelial layer (endosteum) at inner bone surface. The center of the bone marrow is formed by endothelially (en) lined capillaries (cp) and sinusoideal veins (sv) that are connected with each other and the bone surface by a meshwork of reticular cells (rc; green). Reticular cells form the stroma of the bone marrow, which contains proliferating and differentiating clusters of hematopoietic stem cells (HSCs) (denoted in figure by hsc; violet) and blood progenitors (bp; brown) as well as postmitotic maturing blood cells. HSCs are enriched in the zone that contacts osteoblasts; this zone is believed to constitute the HSC niche. Differentiating blood cells are seen to enter the circulation; these include thrombocytes (tc), produced by the fragmentation of large megakaryocytes (mgc), and leukocytes (lc).
(c) Histological section of human bone marrow showing the interface of bone matrix (cb), covered by osteoblasts (ob), with HSCs and blood progenitors (bp) (from Balduino et al. 2005, with permission).
(d ) Scanning electron micrograph of inner bone surface with osteoblasts (ob) and HSCs (from Balduino et al. 2005, with permission).
(e, f ) Confocal section of embryonic mouse bone marrow. Trabecle of bone (cb) is surrounded by osteoblasts (ob) and bone marrow (blue; global nuclear labeling). Panels e and f from Zhang et al. 2003, with permission. (e) HSCs are labeled in pink by stem cell marker Sca; the green label represents BrdU-positive proliferating cells.
( f ) Contact of osteoblasts (ob), marked by expression of N-cadherin ( yellow), with HSCs (pink; labeled with BrdU).
Endothelial cells, osteoblasts, and stromal (also called reticular) cells that crisscross the space between vessels and endosteum form a threedimensional scaffold that houses clusters of blood-forming cells. This scaffold provides a complex microenvironment that, by means of cell-membrane-bound and secreted factors, controls the determination and proliferation of the different blood cell lineages. The multipotent hematopoietic stem cells (HSCs) that seed the bone marrow in the late embryo settle at the outer (subendosteal) layer in contact with the osteoblasts (Figure 5d–f ); these latter cells, forming the stem cell niche of the bone marrow, emit signals that maintain HSCs in their noncommitted stem cell mode. HSC-derived cells that lose contact with the osteoblast layer progress toward the next stage, that of a committed progenitor for lymphoid cells, red blood cells, thrombocytes, (neutrophile) granulocyte/monocytes, basophile granulocytes, or eosinophile granulocytes. These different progenitors are then found nearer the center of the bone marrow, where they proliferate and form growing colonies of maturing blood cells (Figure 5b,c). Once matured, blood cells become capable of crossing the endothelium into the bloodstream. Lymphoid progenitors leave the bone marrow at an immature state and populate the thymus and lymphoid organs.
Hematopoiesis and Hematopoietic Organs in Arthropods 1
Most animals possess populations of cells that are not fixed in place as part of an organ, but that roam within the body cavities to fulfill functions involved with the immune response, cell degradation and replacement, and the distribution of nutrients and gases. The blood cells, or hemocytes, of the vertebrate body, which comprise many different classes each responsible for one of the functions mentioned above, represent the best known example. All invertebrates which possess a body cavity and, to various degrees, a vascular system also have hemocytes of different types. Even in many animals without a body cavity, including cnidaria, sponges and flatworms, one observes motile cells that primarily function as pools of pluripotent stem cells. Such cells may represent the ancestral type of motile cell from which the diverse, specialized classes of hemocytes of higher animals evolved (van de Vyver, 1981).
Hemocytes typically have a lifespan of days to weeks, and need to be replaced constantly. Furthermore, given that the demand for hemocytes (in particular those involved in the immune response, and the related processes of wound healing and regeneration) varies over time, the rate at which hemocytes are produced must be able to adapt to these variable conditions. Blood cells are produced in specialized tissues, called hematopoietic organs. Hematopoietic organs generate blood cells in high numbers throughout the life time of the animal, implying that hematopoietic organs must contain, or be supplied with, self renewing stem cells. The study of hematopoiesis thus addresses a central issue of stem cell biology, namely the “decision” of an undifferentiated cell to either continue to proliferate, or to leave this state to initiate differentiation. We will in the following start out by briefly summarizing pertinent facts about blood cell development among the invertebrates and then provide more details illustrating the architecture and proliferative function of hematopoietic organs in the arthropods.
Hematopoietic cells are part of the vasculature or derive from common blood-vascular progenitors (hemangioblasts)
Hematopoietic organs are built as more or less complex, mesenchymal or gland-like structures that are closely associated with the epithelial cells surrounding blood vessels (endothelia) and/or the coelomic cavity (mesothelial or peritoneal cells), which reflects the shared developmental origin of the motile blood cells and the fixed epithelial cells lining the cavities in which blood cells move. In many phyla, cells scattered randomly all over the endothelia/mesothelia, or located in dedicated hematopoietic “niches” within these epithelia, show higher rates of proliferation and bud off hemocytes into the lumen of the coelom or blood vessels (Fig.1A, B; Eckelbarger, 1976; reviewed in Dales and Dixon, 1981).Origin of hematopoietic tissues.
A: In many coelomate invertebrates (such as polychaete annelids represented here as schematic cross section) hemocytes are produced in hematopoietic centers within the somatopleura, splanchnopleura, and blood vessel wall
B: Histological section of somatopleura of the polychaete Nicolea zostericola, showing hematopoietic center from where groups of hemocytes bud off into the coelom (Scale bar: 18 m; from Eckelbarger, 1976, with permission).
C: Schematic cross section of insect showing hematopoietic lymph gland (lg) closely associated with the dorsal blood vessel.
D: Histological section of the compact hematopoietic organ (lymph gland) of lepidopteran Manduca sexta (Scale bar: 50 m; from Nardi et al., 2003, with permission).
In these cases, progenitors of hemocytes and endothelial/mesothelial cells form one and the same cell type. In other cases, hematopoietic cells form mesenchymal clusters (“nodules”) or gland-like structures (“lymph glands”) that are separate from the vascular linings, but always remain in close contact with them (Fig.1C, D). In Drosophila, where the development of an invertebrate hematopoietic organ was studied, it became clear that a common progenitor (hemangioblast) gives rise to both vascular cells and hemocytes (Mandal et al., 2004), which is similar to the situation in vertebrate embryos (Choi et al., 1998; Teixera et al., 2011). In the Drosophila embryo, vascular progenitors undergo a mesenchymal-epithelial transition to form the lining of the blood vessels; blood progenitors assemble into mesenchymal clusters that remain attached to the basal surface of the vascular epithelia. These clusters proliferate rapidly and develop into the lymph glands of the larva. Lymph glands have been observed in many insects. They form metamerically reiterated encapsulated structures that flank the wall of the dorsal blood vessel (Cuenot, 1897; Hoffmann et al., 1979, 1979). Similar structures have also been described for mollusks (e.g., the amebocyte producing organ in gastropods; Jeong et al., 1983; the “white glands” of cephalopods; Cowden and Curtis, 1981) and crustaceans (lymph glands along the vessels and over the foregut; reviewed inBauchau, 1981). In lower deuterostomes (e.g., ascidians), “blood forming nodules” are grouped around the transverse bars of the pharyngeal basket (Wright, 1981).
Blood Cell Types in Arthropods
Hemocytes have been studied in a number of insects, crustaceans and chelicerates, with much controversy associated with the number of different hemocyte cell types observed and the terminology used to designate them. Thus, often researchers used different terms for the same type of cell; or, differences in cell structure that lead to different names merely reflected artefacts of fixation, or different stages during the development of the same cell. Attempts to simplify and unify the terminology have been made repeatedly. The more recent literature recognizes the following main cell types (reviewed in Jones, 1970; Rowley and Ratcliffe, 1981; Pelc, 1986; Lavine and Strand 2002; Ribeiro and Brehelin, 2006): prohemocytes, plasmatocytes, granulocytes, oenocytoids, and spherule cells (also called adipohemocytes). (1) Prohemocytes (Fig.2D) are small, rounded cells with a high nucleus:cytoplasm ratio and no specialized cytoplasmic organelles; these cells represent the immature hemocyte progenitors, or stem cells. All other blood cell types are derived from prohemocytes. (2) Plasmatocytes (Fig.2A) have more cytoplasm than prohemocytes, and their membrane forms ruffles and filopodia. Plasmatocytes are phagocytotically active, ingesting cellular debris during normal development, and engulfing microorganisms during the immune response. Plasmatocytes are also able to differentiate into large, flattened cells that arrange themselves around foreign bodies that have gained access to the body cavity, forming a capsule around these invaders (Fig.2J). These enlarged, flattened plasmatocytes were called lamellocytes in Drosophila and other insects (Rowley and Ratcliffe, 1981). (3) Granulocytes (Fig.2B) have electron-dense, regularly sized, acidophilic granula. Like plasmatocytes, granulocytes also act as phagocytes. Some insects, including Drosophila, lack granulocytes; here, plasmatocytes take on the function of granulocytes. The loss of granulocytes is apparently not evolutionary ancient; other Dipterans, such as Calliphora, possess granulocytes (Kaaya and Ratcliffe, 1982). (4) Oenocytoids (Fig.2E) are large, oval cells whose cytoplasm contains agglomerates of crystalline material and/or microtubules. The so called crystal cells of Drosophila (Fig.2C) correspond to the oenocytoids defined for other arthropods (Brehelin, 1982). Oenocytoids store prophenol oxidase, an enzyme that plays an important role in the innate immune response (see below). (5) Spherulocytes (Fig.2F) or adipohemocytes are blood cells with basophilic, variably sized and shaped granules. These cells may represent late stages in the differentiative pathway of phagocytic plasmatocytes. The range of blood cells characterized above is simpler, or modified, in some cases. For example, in Drosophila, we do not encounter granulocytes or spherulocytes. Crustaceans and many chelicerates lack oenocytoids; prophenol oxidase is produced by granulocytes (Zhang et al., 2006; Lin and Soderhall, 2011).
A-F: Types of arthropod hemocytes (summarized in E), including
(A) plasmatocytes (pl)(from Rowley and Ratcliffe, 1981, with permission),
(B) granulocytes (gr) (from Rowley and Ratcliffe, 1981, with permission),
(C) oenocytoids/crystal cells (oe/cr)(from Rizki and Rizki, 1984, with permission),
(D) prohemocytes (pr)(from Hoffmann et al., 1979, with permission), and
(F) spherulocytes/adipohemocytes (sp/ad) (from Rowley and Ratcliffe, 1981, with permission).
G-J: Schematic depiction of innate immune reaction carried out by different classes of hemocytes in response to invading microorganisms (ba bacteria; from Rowley and Ratcliffe, 1981, with permission). For detail, see text. Scale bar (A-D, F): 2μm.
Arthropod hemocytes mainly function during the innate immune response and wound healing (Rowley and Ratcliffe, 1981; Nappi et al., 2009; Jiang et al., 2010; Krzemien et al., 2010; Lin and Soderhall, 2011). Invading microorganisms attract plasmatocytes and granulocytes, which release reagents that entrap the invaders (Fig.2G, H). Products secreted by granulocytes also play a role in hemolymph coagulation. Plasmatocytes, differentiating as lamellocytes, form a capsule around the invaders (Fig.2I, J). Oenocytes release prophenol oxidase, which is converted into activated phenol oxidase by a cascade of enzymes that form part of the hemolymph, and that are activated by foreign molecules (e.g., microbial lipopolysaccharides). Phenoloxidase catalyzes a reaction by which melanin is synthesized (Fig.2H-J), and toxic metabolites (e.g., quinones) are released. Encapsulation and melanization act to neutralize the invading microbes or parasites.
How is bug blood different from our own? 2
The major difference between insect blood and the blood of vertebrates, including humans, is that vertebrate blood contains red blood cells. Insects and other invertebrates, on the other hand, have what is called hemolympha heterogeneous fluid that courses through their bodies, bathing all the internal tissues. Hemolymph is mostly water, but it also contains ions, carbohydrates, lipids, glycerol, amino acids, hormones, some cells and pigments. The pigments, however, are usually rather bland, and thus insect blood is clear or tinged with yellow or green. (The red color that you see upon squashing a housefly or fruit fly is actually pigment from the animal's eyes.)
Unlike the closed circulatory system found in vertebrates, insects have an open system lacking arteries and veins. The hemolymph thus flows freely throughout their bodies, lubricating tissues and transporting nutrients and wastes. Whereas the vertebrate circulatory system serves primarily to carry oxygen throughout the body, insects respire an entirely different waynamely, through tracheal tubes. In the case of the fruit fly Drosophila, for example, a series of tiny openings called spiracles line the impermeable outer skin of the fly, and these convey air directly to tracheal tubes that, in turn, convey air to the internal tissues.
Insects do have hearts that pump the hemolymph throughout their circulatory systems. Though these hearts are quite different from vertebrate hearts, some of the genes that direct heart development in the two groups are in fact very similar. The development and evolution of the vertebrate heart is currently the subject of much research.
1) http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3873168/
2) http://www.scientificamerican.com/article/how-is-bug-blood-differen/
3) http://materiais.dbio.uevora.pt/BD/Diferenciacao/Hemacytoblast.pdf
4) ( Mesenchymal stem cells, or MSCs, are multipotent stromal cells that can differentiate into a variety of cell types )
5) ( Refers to the main body cavity in most multicellular animals .A body cavity is any fluid-filled space in a multicellular organism other than those of vessels (such as blood vessels and lymph vessels). The term usually refers to the space located between an animal’s outer covering (epidermis), and the outer lining of the gut cavity, where internal organs developIn the simplest scenario (e.g., some polychaetes)
6) The mesothelium is a membrane composed of simple squamous cells that forms the lining of several body cavities: the pleura (thoracic cavity), peritoneum (abdominal cavity including the mesentery), mediastinum andpericardium (heart sac). Mesothelial tissue also surrounds the male internal reproductive organs (the tunica vaginalis testis) and covers the internal reproductive organs of women (the tunica serosa uteri). Mesothelium that covers the internal organs is called visceral mesothelium, while the layer that covers the body walls is called the parietal mesothelium. Mesothelium is the epithelial component of serosa.
7) Deuterostomes (taxonomic term: Deuterostomia; from the Greek: "mouth second") are any members of a superphylum of animals. Deuterostomia is a subtaxon of the Bilateria branch of the subkingdom Eumetazoa, within Animalia, and are distinguished from protostomes by their embryonic development; in deuterostomes, the first opening (the blastopore) becomes the anus, while in protostomes, it becomes the mouth. Deuterostomes are also known as enterocoelomates because their coelom develops through enterocoely.
8 Mesenchymal stem cells, or MSCs, are multipotent stromal cells that can differentiate into a variety of cell types, including: osteoblasts (bone cells), chondrocytes (cartilage cells), myocytes (muscle cells) andadipocytes (fat cells). This phenomenon has been documented in specific cells and tissues in living animals and their counterparts growing in tissue culture.