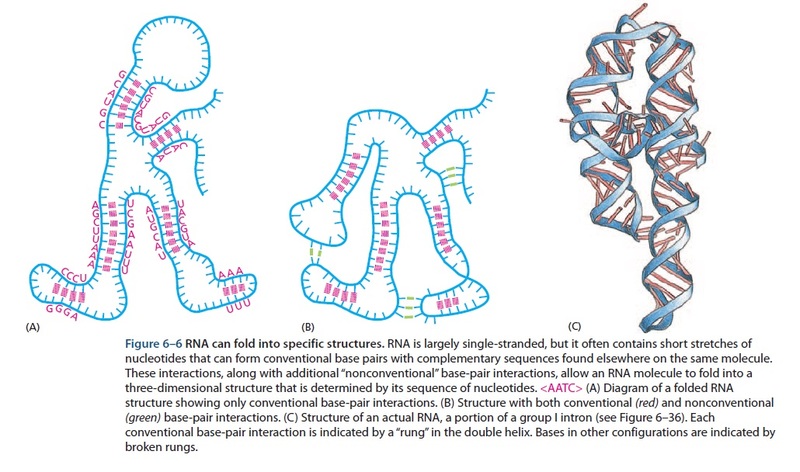
No evidence that RNA molecules ever had the broad range of catalytic activities
https://reasonandscience.catsboard.com/t2243-no-evidence-that-rna-molecules-ever-had-the-broad-range-of-catalytic-activities
Paul Davies The Algorithmic Origins of Life
Despite the conceptual elegance of the RNA world, the hypothesis faces problems, primarily due to the immense challenge of synthesizing RNA nucleotides under plausible prebiotic conditions and the susceptibility of RNA oligomers to degradation via hydrolysis 21 Due to the organizational structure of systems capable of processing algorithmic (instructional) information, it is not at all clear that a monomolecular system – where a single polymer plays the role of catalyst and informational carrier – is even logically consistent with the organization of information flow in living systems, because there is no possibility of separating information storage from information processing (that being such a distinctive feature of modern life). As such, digital–first systems (as currently posed) represent a rather trivial form of information processing that fails to capture the logical structure of life as we know it.
We need to explain the origin of both the hardware and software aspects of life, or the job is only half finished. Explaining the chemical substrate of life and claiming it as a solution to life’s origin is like pointing to silicon and copper as an explanation for the goings-on inside a computer. It is this transition where one should expect to see a chemical system literally take-on “a life of its own”, characterized by informational dynamics which become decoupled from the dictates of local chemistry alone (while of course remaining fully consistent with those dictates). Thus the famed chicken-or-egg problem (a solely hardware issue) is not the true sticking point. Rather, the puzzle lies with something fundamentally different, a problem of causal organization having to do with the separation of informational and mechanical aspects into parallel causal narratives. The real challenge of life’s origin is thus to explain how instructional information control systems emerge naturally and spontaneously from mere molecular dynamics.
Systems of interconnected software and hardware like in the cell are irreducibly complex and interdependent. There is no reason for information processing machinery to exist without the software, and vice versa.
Perry Marshall, Evolution 2.0, page 151:
There are problems with the RNA world hypothesis: Many scientists believe RNA is too complex to have arisen without the presence of the very same life forms it is believed to have created; RNA is inherently unstable, so even if it did arise, it wouldn’t last long without a cell to protect it; catalysis of chemical reactions is seldom observed to occur in long RNA sequences only; and the catalytic abilities of RNA are limited. The RNA world hypothesis doesn’t actually solve the chicken-and-egg problem of RNA and proteins: You need RNA to produce proteins, but you need proteins to build the machinery to read the RNA in the first place. As a communication engineer, my objection to the RNA hypothesis is that to evolve any kind of cell, RNA would have to self-replicate. But the RNA strand formation you read about in the literature is not codebased self-replication. It’s similar to crystal growth, which does not use codes at all! RNA strand formation in a chemical lab is not in any way, shape, or form the same as DNA transcription and translation. In DNA transcription and translation, in order to convert code to proteins, you need a ribosome to transcribe the message. But in order to have a ribosome you have to have a plan for building a ribosome first. A ribosome is partly made from RNA. So before that, you have to have a code in the RNA. Many books and papers on the Origin of Life only discuss the assembly of the chemicals themselves. Nothing we know about chemicals tells us where codes come from. Saying you can get real DNA by stringing chemicals together is like telling your kid that TVs come from a glass factory
Without code there can be no self-replication. Without self-replication you can’t have reproduction. Without reproduction you can’t have evolution or natural selection.
from the book: The Logic of Chance: The Nature and Origin of Biological Evolution
By Eugene V. Koonin
https://reasonandscience.catsboard.com/t2234-the-origin-of-replication-and-translation-and-the-rna-world
The origin of replication and translation and the RNA World
The primary incentive behind the theory of self-replicating systems that Manfred Eigen outlined was to develop a simple model explaining the origin of biological information and, hence, of life itself. Eigen’s theory revealed the existence of the fundamental limit on the fidelity of replication (the Eigen threshold): If the product of the error (mutation) rate and the information capacity (genome size) is below the Eigen threshold, there will be stable inheritance and hence evolution; however, if it is above the threshold, the mutational meltdown and extinction become inevitable (Eigen, 1971). The Eigen threshold lies somewhere between 1 and 10 mutations per round of replication (Tejero, et al., 2011); regardless of the exact value, staying above the threshold fidelity is required for sustainable replication and so is a prerequisite for the start of biological evolution
Indeed, the very origin of the first organisms presents at least an appearance of a paradox because a certain minimum level of complexity is required to make self-replication possible at all; high-fidelity replication requires additional functionalities that need even more information to be encoded (Penny, 2005). However, the replication fidelity at a given point in time limits the amount of information that can be encoded in the genome. What turns this seemingly vicious circle into the (seemingly) unending spiral of increasing complexity—the Darwin-Eigen cycle, following the terminology introduced by David Penny (Penny, 2005)—is a combination of natural selection with genetic drift. Even small gains in replication fidelity are advantageous to the system, if only because of the decrease of the reproduction cost as a result of the increasing yield of viable copies of the genome. In itself, a larger genome is more of a liability than an advantage because of higher replication costs. However, moderate genome increase, such as by duplication of parts of the genome or by recombination, can be fixed via genetic drift in small populations. Replicators with a sufficiently high fidelity can take advantage of such randomly fixed and initially useless genetic material by evolving new functions, without falling off the “Eigen cliff” (see Figure 12-1B). Among such newly evolved, fitness-increasing functions will be those that increase replication fidelity, which, in turn, allows a further increase in the amount of encoded information. And so the Darwin- Eigen cycle recapitulates itself in a spiral progression, leading to a steady increase in genome complexity (see Figure 12-1A). The crucial question in the study of the origin of life is how the Darwin-Eigen cycle started—how was the minimum complexity that is required to achieve the minimally acceptable replication fidelity attained? In even the simplest modern systems, such as RNA viruses with the replication fidelity of only about 10^3 and viroids that replicate with the lowest fidelity among the known replicons (about 10^2; Gago, et al., 2009), replication is catalyzed by complex protein polymerases. The replicase itself is produced by translation of the respective mRNA(s), which is mediated by the immensely complex ribosomal apparatus. Hence, the dramatic paradox of the origin of life is that, to attain the minimum complexity required for a biological system to start on the Darwin-Eigen spiral, a system of a far greater complexity appears to be required. How such a system could evolve is a puzzle that defeats conventional evolutionary thinking, all of which is about biological systems moving along the spiral; the solution is bound to be unusual.
Or the solution might be outside the realm of philosophical naturalism, that is, intelligent design ?!!
A protein must be able to fold into a specific 3-dimensional shape in order to have biological activity. But the forces holding the folded protein in shape are so weak that many amino acids need to be involved - imposing a minimum length on their sequence of about 70 (Kyte), and maybe 50 for nucleic acids. So trying to improve the odds of finding a biologically active macromolecule by starting with short ones, just will not work.
A similar misperception is that the first replicator need only have had poor replicating ability, which could gradually have improved (by mutation and selection of improved versions). But it is important to note that a poor replicator is more likely to degrade through miscopying than to improve its performance, and this poses a dilemma for the production of a primitive replicator. Although the common presumption is that a crude replicator can gradually improve its performance through a natural selection sort of process, in fact there is a threshold before that could take place. That is, a replicator must already have a reasonably good performance in order to be able to improve on that performance.
In other words, natural selection cannot take place until there is a reasonably reliable replicating system. So the first replicating system would need to have arisen exclusively by chance.
The "RNA World" is essentially a hypothetical stage of life between the first replicating molecule and the highly complicated DNA-protein-based life. The chief problem facing an RNA world is that RNA cannot perform all of the functions of DNA adequately to allow for replication and transcription of proteins.
New findings challenge assumptions about origins of life
https://reasonandscience.catsboard.com/t1428-replicator-first-and-metabolism-first-scenarios
There is currently no known chemical pathway for an "RNA world" to transform into a "DNA/protein world."
http://phys.org/news/2013-09-assumptions-life.html#jCp
But for the hypothesis to be correct, ancient RNA catalysts would have had to copy multiple sets of RNA blueprints nearly as accurately as do modern-day enzymes. That's a hard sell; scientists calculate that it would take much longer than the age of the universe for randomly generated RNA molecules to evolve sufficiently to achieve the modern level of sophistication. Given Earth's age of 4.5 billion years, living systems run entirely by RNA could not have reproduced and evolved either fast or accurately enough to give rise to the vast biological complexity on Earth today.
OOL theorist Leslie Orgel notes that an "RNA World" could only form the basis for life, "if prebiotic RNA had two properties not evident today: a capacity to replicate without the help of proteins and an ability to catalyze every step of protein synthesis." The RNA world is thus a hypothetical system behind which there is little positive evidence, and much materialist philosophy: "The precise events giving rise to the RNA world remain unclear … investigators have proposed many hypotheses, but evidence in favor of each of them is fragmentary at best. The full details of how the RNA world, and life, emerged may not be revealed in the near future.
http://www.ncbi.nlm.nih.gov/pubmed/22793875
(i) RNA is too complex a molecule to have arisen prebiotically;
(ii) RNA is inherently unstable;
(iii) catalysis is a relatively rare property of long RNA sequences only; and
(iv) the catalytic repertoire of RNA is too limited.
The best claimed evidence of an "RNA World" includes the fact that there are RNA enzymes and genomes, and that cells use RNA to convert the DNA code into proteins. However, RNA plays only a supporting role in the cell, and there is no known biochemical system completely composed of RNA.
RNA experts have created a variety of RNA molecules which can perform biochemical functions through what is commonly termed "test tube evolution." However, "test tube evolution" is just a description for what is in reality nothing more than chemical engineering in the laboratory employing Darwinian principles; that does not imply that there is some known pathway through which these molecules could arise naturally.
In order a molecule to be a self replicator , it has to be a homopolymer, of which the backbone must have the same repetitive units; they must be identical. On the prebiotic world, the generation of a homopolymer was however impossible.
Steven A. Benner, Ph.D. Chemistry, Harvard, prominent origin-of-life researcher and creator of the Foundation for Applied Molecular Evolution, was posted on Huffington Post on December 6, 2013. In it he said,
"We have failed in any continuous way to provide a recipe that gets from the simple molecules that we know were present on early Earth to RNA."
That lead Leslie Orgel to say :
It would take a miracle if a strand of RNA ever appeared on the primitive Earth.
(Dover, 1999, p. 218).
I would have thought it relevant to point out for biologists in general that not one self-replicating RNA has emerged to date from quadrillions (1024) of artificially synthesized, random RNA sequences
How could the first living cells with DNA-based molecular biology have originated by spontaneous chemical processes on the prebiotic Earth? Primordial DNA synthesis would have required the presence of specific enzymes, but how could these enzymes be synthesized without the genetic information in DNA and without RNA for translating that information into the amino acid sequence of the protein enzymes? In other words, proteins are required for DNA synthesis and DNA is required for protein synthesis.
This classic "chicken-and-egg" problem made it immensely difficult to conceive of any plausible prebiotic chemical pathway to the molecular biological system. Certainly no such chemical pathway had been demonstrated 2
Joyce and Orgel note, it seems unlikely that a structure with fewer than 40 nucleotides would be sufficient. Suppose, then, that "there is some 50-mer [RNA molecule of 50 nucleotides length]," Joyce and Orgel speculate, that "replicates with 90% fidelity. ... Would such a molecule be expected to occur within a population of random RNAs?"
Perhaps: but one such self-replicating molecule will not suffice.
"Unless the molecule can literally copy itself," Joyce and Orgel note, "that is, act simultaneously as both template and catalyst, it must encounter another copy of itself that it can use as a template." Copying any given RNA in its vicinity will lead to an error catastrophe, as the population of RNAs will decay into a collection of random sequences. But to find another copy of itself, the self-replicating RNA would need (Joyce and Orgel calculate) a library of RNA that "far exceeds the mass of the earth."18
In the face of these difficulties, they advise, one must reject the myth of a self-replicating RNA molecule that arose de novo from a soup of random polynucleotides. Not only is such a notion unrealistic in light of our current understanding of prebiotic chemistry, but it should strain the credulity of even an optimist's view of RNA's catalytic potential. If you doubt this, ask yourself whether you believe that a replicase ribozyme would arise in a solution containing nucleoside 5'-diphosphates and polynucleotide phosphorylase!
G. F. Joyce, L. E. Orgel, "Prospects for Understanding the Origin of the RNA World," In the RNA World, Cold Spring Harbor Laboratory Press, New York, 1993, p. 13.
This discussion… has, in a sense, focused on a straw man: the myth of a self-replicating RNA molecule that arose de novo from a soup of random polynucleotides. Not only is such a notion unrealistic in light of our current understanding of prebiotic chemistry, but it would strain the credulity of even an optimist's view of RNA's catalytic potential
Even if we suppose that there was self-replicating RNA in the primordial world, that numerous amino acids of every type ready to be used by RNA were available, and that all of these impossibilities somehow took place, the situation still does not lead to the formation of even one single protein. For RNA only includes information concerning the structure of proteins. Amino acids, on the other hand, are raw materials. Nevertheless, there is no mechanism for the production of proteins. To consider the existence of RNA sufficient for protein production is as nonsensical as expecting a car to assemble itself by simply throwing the blueprint onto a heap of parts piled up on top of each other. A blueprint cannot produce a car all by itself without a factory and workers to assemble the parts according to the instructions contained in the blueprint; in the same way, the blueprint contained in RNA cannot produce proteins by itself without the cooperation of other cellular components which follow the instructions contained in the RNA.
The problem of the origin of the RNA World is far from being solved. One can sketch out a logical order of events, beginning with prebiotic chemistry and ending with DNA/protein-based life. However, it must be said that the details of this process remain obscure and are not likely to be known in the near future. 3
A cell has a great investment in its RNAs – they are working copies of its genomic information. The study of mRNA biogenesis in the last few years has revealed an elaborate surveillance mechanism involving factors such as the UPF proteins that culls aberrantly spliced mRNAs and mRNAs with premature termination codons. There might be a hint that such RNA quality control mechanisms go awry in cancers, just as DNA quality control mechanisms do, where aberrantly spliced transcripts accumulate in a tumor. Now that the gates are open, we may have a flood of studies on the RNome [the RNA genome] stability and cancer. 1
This aggravates the chicken-and-egg problem for proponents of natural mechanisms. In the “RNA World” hypothesis for the origin of life, RNA performed both the information storage and enzymatic functions before these roles were outsourced to DNA and proteins. But how could RNA repair itself? If RNA needs to be protected from damage, the protein repair system would have needed to be there from the beginning. Proponents of natural mechanisms might surmise that different primitive RNAs worked side by side to repair each other, but that strains credibility for a hypothesis already far-fetched.
In typical evolutionary lingo, Begley and Samson blow smoke about what nature produced (emphasis added): “It seems that, for each human protein, parameters have evolved to distinguish between RNA and DNA,” they speculate, and in another place, “It might be that the RNA-demethylation activity of AlkB-like proteins evolved to regulate biological RNA methylation, and that the repair of aberrant, chemical methylation is fortuitous.” Ask them how the cell evolved these things, and you’ll probably get a quizzical look, as if “Why are you asking such a dumb question? I don’t know. It just had to. We’re here, aren’t we?”1) http://reasonandscience.heavenforum.org/t2043-dna-and-rna-error-checking-and-repair-amazing-evidence-of-design#4671
The Logic of Chance: The Nature and Origin of Biological Evolution, Eugene V. Koonin, page 365:
The RNA World hypothesis faces grave difficulties. First, despite all invested effort, the in vitro evolved ribozymes remain (relatively) poor catalysts for most reactions; the lack of efficient, processive ribozyme polymerases seems particularly troubling, but there is also a serious shortage of other activities, such as those required for the synthesis of nucleotides. Admittedly, it might be unrealistic to expect that experiments on in vitro evolution of ribozymes could easily mimic the actual complexity of the primordial RNA World. Although these experiments harness the power of selection, they are obviously performed on a totally different time scale and under conditions that cannot accurately reproduce the (unknown) conditions at the origin of life
The overall situation in the origin of life field appears rather grim. Even under the (highly nontrivial) assumption that monomers such as NTP are readily available, the problem of the synthesis of sufficiently stable, structurally regular polymers (RNA) is formidable, and the origin of replication and translation from such primordial RNA molecules could be an even harder problem. As emphasized repeatedly in this book, evolution by natural selection
and drift can begin only after replication with sufficient fidelity is established. Even at that stage, the evolution of translation remains highly problematic.
https://reasonandscience.catsboard.com/t2243-no-evidence-that-rna-molecules-ever-had-the-broad-range-of-catalytic-activities
Paul Davies The Algorithmic Origins of Life
Despite the conceptual elegance of the RNA world, the hypothesis faces problems, primarily due to the immense challenge of synthesizing RNA nucleotides under plausible prebiotic conditions and the susceptibility of RNA oligomers to degradation via hydrolysis 21 Due to the organizational structure of systems capable of processing algorithmic (instructional) information, it is not at all clear that a monomolecular system – where a single polymer plays the role of catalyst and informational carrier – is even logically consistent with the organization of information flow in living systems, because there is no possibility of separating information storage from information processing (that being such a distinctive feature of modern life). As such, digital–first systems (as currently posed) represent a rather trivial form of information processing that fails to capture the logical structure of life as we know it.
We need to explain the origin of both the hardware and software aspects of life, or the job is only half finished. Explaining the chemical substrate of life and claiming it as a solution to life’s origin is like pointing to silicon and copper as an explanation for the goings-on inside a computer. It is this transition where one should expect to see a chemical system literally take-on “a life of its own”, characterized by informational dynamics which become decoupled from the dictates of local chemistry alone (while of course remaining fully consistent with those dictates). Thus the famed chicken-or-egg problem (a solely hardware issue) is not the true sticking point. Rather, the puzzle lies with something fundamentally different, a problem of causal organization having to do with the separation of informational and mechanical aspects into parallel causal narratives. The real challenge of life’s origin is thus to explain how instructional information control systems emerge naturally and spontaneously from mere molecular dynamics.
Systems of interconnected software and hardware like in the cell are irreducibly complex and interdependent. There is no reason for information processing machinery to exist without the software, and vice versa.
Perry Marshall, Evolution 2.0, page 151:
There are problems with the RNA world hypothesis: Many scientists believe RNA is too complex to have arisen without the presence of the very same life forms it is believed to have created; RNA is inherently unstable, so even if it did arise, it wouldn’t last long without a cell to protect it; catalysis of chemical reactions is seldom observed to occur in long RNA sequences only; and the catalytic abilities of RNA are limited. The RNA world hypothesis doesn’t actually solve the chicken-and-egg problem of RNA and proteins: You need RNA to produce proteins, but you need proteins to build the machinery to read the RNA in the first place. As a communication engineer, my objection to the RNA hypothesis is that to evolve any kind of cell, RNA would have to self-replicate. But the RNA strand formation you read about in the literature is not codebased self-replication. It’s similar to crystal growth, which does not use codes at all! RNA strand formation in a chemical lab is not in any way, shape, or form the same as DNA transcription and translation. In DNA transcription and translation, in order to convert code to proteins, you need a ribosome to transcribe the message. But in order to have a ribosome you have to have a plan for building a ribosome first. A ribosome is partly made from RNA. So before that, you have to have a code in the RNA. Many books and papers on the Origin of Life only discuss the assembly of the chemicals themselves. Nothing we know about chemicals tells us where codes come from. Saying you can get real DNA by stringing chemicals together is like telling your kid that TVs come from a glass factory
Without code there can be no self-replication. Without self-replication you can’t have reproduction. Without reproduction you can’t have evolution or natural selection.
from the book: The Logic of Chance: The Nature and Origin of Biological Evolution
By Eugene V. Koonin
https://reasonandscience.catsboard.com/t2234-the-origin-of-replication-and-translation-and-the-rna-world
The origin of replication and translation and the RNA World
The primary incentive behind the theory of self-replicating systems that Manfred Eigen outlined was to develop a simple model explaining the origin of biological information and, hence, of life itself. Eigen’s theory revealed the existence of the fundamental limit on the fidelity of replication (the Eigen threshold): If the product of the error (mutation) rate and the information capacity (genome size) is below the Eigen threshold, there will be stable inheritance and hence evolution; however, if it is above the threshold, the mutational meltdown and extinction become inevitable (Eigen, 1971). The Eigen threshold lies somewhere between 1 and 10 mutations per round of replication (Tejero, et al., 2011); regardless of the exact value, staying above the threshold fidelity is required for sustainable replication and so is a prerequisite for the start of biological evolution
Indeed, the very origin of the first organisms presents at least an appearance of a paradox because a certain minimum level of complexity is required to make self-replication possible at all; high-fidelity replication requires additional functionalities that need even more information to be encoded (Penny, 2005). However, the replication fidelity at a given point in time limits the amount of information that can be encoded in the genome. What turns this seemingly vicious circle into the (seemingly) unending spiral of increasing complexity—the Darwin-Eigen cycle, following the terminology introduced by David Penny (Penny, 2005)—is a combination of natural selection with genetic drift. Even small gains in replication fidelity are advantageous to the system, if only because of the decrease of the reproduction cost as a result of the increasing yield of viable copies of the genome. In itself, a larger genome is more of a liability than an advantage because of higher replication costs. However, moderate genome increase, such as by duplication of parts of the genome or by recombination, can be fixed via genetic drift in small populations. Replicators with a sufficiently high fidelity can take advantage of such randomly fixed and initially useless genetic material by evolving new functions, without falling off the “Eigen cliff” (see Figure 12-1B). Among such newly evolved, fitness-increasing functions will be those that increase replication fidelity, which, in turn, allows a further increase in the amount of encoded information. And so the Darwin- Eigen cycle recapitulates itself in a spiral progression, leading to a steady increase in genome complexity (see Figure 12-1A). The crucial question in the study of the origin of life is how the Darwin-Eigen cycle started—how was the minimum complexity that is required to achieve the minimally acceptable replication fidelity attained? In even the simplest modern systems, such as RNA viruses with the replication fidelity of only about 10^3 and viroids that replicate with the lowest fidelity among the known replicons (about 10^2; Gago, et al., 2009), replication is catalyzed by complex protein polymerases. The replicase itself is produced by translation of the respective mRNA(s), which is mediated by the immensely complex ribosomal apparatus. Hence, the dramatic paradox of the origin of life is that, to attain the minimum complexity required for a biological system to start on the Darwin-Eigen spiral, a system of a far greater complexity appears to be required. How such a system could evolve is a puzzle that defeats conventional evolutionary thinking, all of which is about biological systems moving along the spiral; the solution is bound to be unusual.
Or the solution might be outside the realm of philosophical naturalism, that is, intelligent design ?!!
A protein must be able to fold into a specific 3-dimensional shape in order to have biological activity. But the forces holding the folded protein in shape are so weak that many amino acids need to be involved - imposing a minimum length on their sequence of about 70 (Kyte), and maybe 50 for nucleic acids. So trying to improve the odds of finding a biologically active macromolecule by starting with short ones, just will not work.
A similar misperception is that the first replicator need only have had poor replicating ability, which could gradually have improved (by mutation and selection of improved versions). But it is important to note that a poor replicator is more likely to degrade through miscopying than to improve its performance, and this poses a dilemma for the production of a primitive replicator. Although the common presumption is that a crude replicator can gradually improve its performance through a natural selection sort of process, in fact there is a threshold before that could take place. That is, a replicator must already have a reasonably good performance in order to be able to improve on that performance.
In other words, natural selection cannot take place until there is a reasonably reliable replicating system. So the first replicating system would need to have arisen exclusively by chance.
The "RNA World" is essentially a hypothetical stage of life between the first replicating molecule and the highly complicated DNA-protein-based life. The chief problem facing an RNA world is that RNA cannot perform all of the functions of DNA adequately to allow for replication and transcription of proteins.
New findings challenge assumptions about origins of life
https://reasonandscience.catsboard.com/t1428-replicator-first-and-metabolism-first-scenarios
There is currently no known chemical pathway for an "RNA world" to transform into a "DNA/protein world."
http://phys.org/news/2013-09-assumptions-life.html#jCp
But for the hypothesis to be correct, ancient RNA catalysts would have had to copy multiple sets of RNA blueprints nearly as accurately as do modern-day enzymes. That's a hard sell; scientists calculate that it would take much longer than the age of the universe for randomly generated RNA molecules to evolve sufficiently to achieve the modern level of sophistication. Given Earth's age of 4.5 billion years, living systems run entirely by RNA could not have reproduced and evolved either fast or accurately enough to give rise to the vast biological complexity on Earth today.
OOL theorist Leslie Orgel notes that an "RNA World" could only form the basis for life, "if prebiotic RNA had two properties not evident today: a capacity to replicate without the help of proteins and an ability to catalyze every step of protein synthesis." The RNA world is thus a hypothetical system behind which there is little positive evidence, and much materialist philosophy: "The precise events giving rise to the RNA world remain unclear … investigators have proposed many hypotheses, but evidence in favor of each of them is fragmentary at best. The full details of how the RNA world, and life, emerged may not be revealed in the near future.
http://www.ncbi.nlm.nih.gov/pubmed/22793875
(i) RNA is too complex a molecule to have arisen prebiotically;
(ii) RNA is inherently unstable;
(iii) catalysis is a relatively rare property of long RNA sequences only; and
(iv) the catalytic repertoire of RNA is too limited.
The best claimed evidence of an "RNA World" includes the fact that there are RNA enzymes and genomes, and that cells use RNA to convert the DNA code into proteins. However, RNA plays only a supporting role in the cell, and there is no known biochemical system completely composed of RNA.
RNA experts have created a variety of RNA molecules which can perform biochemical functions through what is commonly termed "test tube evolution." However, "test tube evolution" is just a description for what is in reality nothing more than chemical engineering in the laboratory employing Darwinian principles; that does not imply that there is some known pathway through which these molecules could arise naturally.
In order a molecule to be a self replicator , it has to be a homopolymer, of which the backbone must have the same repetitive units; they must be identical. On the prebiotic world, the generation of a homopolymer was however impossible.
Steven A. Benner, Ph.D. Chemistry, Harvard, prominent origin-of-life researcher and creator of the Foundation for Applied Molecular Evolution, was posted on Huffington Post on December 6, 2013. In it he said,
"We have failed in any continuous way to provide a recipe that gets from the simple molecules that we know were present on early Earth to RNA."
That lead Leslie Orgel to say :
It would take a miracle if a strand of RNA ever appeared on the primitive Earth.
(Dover, 1999, p. 218).
I would have thought it relevant to point out for biologists in general that not one self-replicating RNA has emerged to date from quadrillions (1024) of artificially synthesized, random RNA sequences
How could the first living cells with DNA-based molecular biology have originated by spontaneous chemical processes on the prebiotic Earth? Primordial DNA synthesis would have required the presence of specific enzymes, but how could these enzymes be synthesized without the genetic information in DNA and without RNA for translating that information into the amino acid sequence of the protein enzymes? In other words, proteins are required for DNA synthesis and DNA is required for protein synthesis.
This classic "chicken-and-egg" problem made it immensely difficult to conceive of any plausible prebiotic chemical pathway to the molecular biological system. Certainly no such chemical pathway had been demonstrated 2
Joyce and Orgel note, it seems unlikely that a structure with fewer than 40 nucleotides would be sufficient. Suppose, then, that "there is some 50-mer [RNA molecule of 50 nucleotides length]," Joyce and Orgel speculate, that "replicates with 90% fidelity. ... Would such a molecule be expected to occur within a population of random RNAs?"
Perhaps: but one such self-replicating molecule will not suffice.
"Unless the molecule can literally copy itself," Joyce and Orgel note, "that is, act simultaneously as both template and catalyst, it must encounter another copy of itself that it can use as a template." Copying any given RNA in its vicinity will lead to an error catastrophe, as the population of RNAs will decay into a collection of random sequences. But to find another copy of itself, the self-replicating RNA would need (Joyce and Orgel calculate) a library of RNA that "far exceeds the mass of the earth."18
In the face of these difficulties, they advise, one must reject the myth of a self-replicating RNA molecule that arose de novo from a soup of random polynucleotides. Not only is such a notion unrealistic in light of our current understanding of prebiotic chemistry, but it should strain the credulity of even an optimist's view of RNA's catalytic potential. If you doubt this, ask yourself whether you believe that a replicase ribozyme would arise in a solution containing nucleoside 5'-diphosphates and polynucleotide phosphorylase!
G. F. Joyce, L. E. Orgel, "Prospects for Understanding the Origin of the RNA World," In the RNA World, Cold Spring Harbor Laboratory Press, New York, 1993, p. 13.
This discussion… has, in a sense, focused on a straw man: the myth of a self-replicating RNA molecule that arose de novo from a soup of random polynucleotides. Not only is such a notion unrealistic in light of our current understanding of prebiotic chemistry, but it would strain the credulity of even an optimist's view of RNA's catalytic potential
Even if we suppose that there was self-replicating RNA in the primordial world, that numerous amino acids of every type ready to be used by RNA were available, and that all of these impossibilities somehow took place, the situation still does not lead to the formation of even one single protein. For RNA only includes information concerning the structure of proteins. Amino acids, on the other hand, are raw materials. Nevertheless, there is no mechanism for the production of proteins. To consider the existence of RNA sufficient for protein production is as nonsensical as expecting a car to assemble itself by simply throwing the blueprint onto a heap of parts piled up on top of each other. A blueprint cannot produce a car all by itself without a factory and workers to assemble the parts according to the instructions contained in the blueprint; in the same way, the blueprint contained in RNA cannot produce proteins by itself without the cooperation of other cellular components which follow the instructions contained in the RNA.
The problem of the origin of the RNA World is far from being solved. One can sketch out a logical order of events, beginning with prebiotic chemistry and ending with DNA/protein-based life. However, it must be said that the details of this process remain obscure and are not likely to be known in the near future. 3
in the early 1980s, the discovery of self-splicing, catalytic RNA molecules (in the ciliated protozoan Tetrahymena thermophila), put molecular flesh on the speculative bones of the idea of an early evolutionary stage dominated by RNA. These catalytic RNA molecules have subsequently been termed "ribozymes." "One can contemplate an RNA World," wrote Walter Gilbert in 1986, "containing only RNA molecules that serve to catalyze the synthesis of themselves."
Micromolecules do not spontaneously combine to form macromolecules.
It is said that DNA is the secret of life. DNA is not the secret of life. Life is the secret of DNA. Evolutionists persistently claim that the initial stage in the origin of life was the origin of a self-replicating DNA or RNA molecule. There is no such thing as a self-replicating molecule, and no such molecule could ever exist.The formation of a molecule requires the input of a highly selected type of energy and the steady input of the building blocks required to form it. To produce a protein, the building blocks are amino acids. For DNA and RNA these building blocks are nucleotides, which are composed of purines, pyrimidines, sugars, and phosphoric acid. If amino acids are dissolved in water they do not spontaneously join together to make a protein. That would require an input of energy. If proteins are dissolved in water the chemical bonds between the amino acids slowly break apart, releasing energy (the protein is said to hydrolyze). The same is true of DNA and RNA. To form a protein in a laboratory the chemist, after dissolving the required amino acids in a solvent, adds a chemical that contains high energy bonds (referred to as a peptide reagent). The energy from this chemical is transferred to the amino acids. This provides the necessary energy to form the chemical bonds between the amino acids and releases H and OH to form H2O (water). This only happens in a chemistry laboratory or in the cells of living organisms. It could never have taken place in a primitive ocean or anywhere on a primitive Earth. Who or what would be there to provide a steady input of the appropriate energy? Destructive raw energy would not work. Who or what would be there to provide a steady supply of the appropriate building blocks rather than just junk? In speaking of a self-replicating DNA molecule, evolutionists are reaching for a pie in the sky.
It is said that DNA is the secret of life. DNA is not the secret of life. Life is the secret of DNA. Evolutionists persistently claim that the initial stage in the origin of life was the origin of a self-replicating DNA or RNA molecule. There is no such thing as a self-replicating molecule, and no such molecule could ever exist.The formation of a molecule requires the input of a highly selected type of energy and the steady input of the building blocks required to form it. To produce a protein, the building blocks are amino acids. For DNA and RNA these building blocks are nucleotides, which are composed of purines, pyrimidines, sugars, and phosphoric acid. If amino acids are dissolved in water they do not spontaneously join together to make a protein. That would require an input of energy. If proteins are dissolved in water the chemical bonds between the amino acids slowly break apart, releasing energy (the protein is said to hydrolyze). The same is true of DNA and RNA. To form a protein in a laboratory the chemist, after dissolving the required amino acids in a solvent, adds a chemical that contains high energy bonds (referred to as a peptide reagent). The energy from this chemical is transferred to the amino acids. This provides the necessary energy to form the chemical bonds between the amino acids and releases H and OH to form H2O (water). This only happens in a chemistry laboratory or in the cells of living organisms. It could never have taken place in a primitive ocean or anywhere on a primitive Earth. Who or what would be there to provide a steady input of the appropriate energy? Destructive raw energy would not work. Who or what would be there to provide a steady supply of the appropriate building blocks rather than just junk? In speaking of a self-replicating DNA molecule, evolutionists are reaching for a pie in the sky.
A cell has a great investment in its RNAs – they are working copies of its genomic information. The study of mRNA biogenesis in the last few years has revealed an elaborate surveillance mechanism involving factors such as the UPF proteins that culls aberrantly spliced mRNAs and mRNAs with premature termination codons. There might be a hint that such RNA quality control mechanisms go awry in cancers, just as DNA quality control mechanisms do, where aberrantly spliced transcripts accumulate in a tumor. Now that the gates are open, we may have a flood of studies on the RNome [the RNA genome] stability and cancer. 1
This aggravates the chicken-and-egg problem for proponents of natural mechanisms. In the “RNA World” hypothesis for the origin of life, RNA performed both the information storage and enzymatic functions before these roles were outsourced to DNA and proteins. But how could RNA repair itself? If RNA needs to be protected from damage, the protein repair system would have needed to be there from the beginning. Proponents of natural mechanisms might surmise that different primitive RNAs worked side by side to repair each other, but that strains credibility for a hypothesis already far-fetched.
In typical evolutionary lingo, Begley and Samson blow smoke about what nature produced (emphasis added): “It seems that, for each human protein, parameters have evolved to distinguish between RNA and DNA,” they speculate, and in another place, “It might be that the RNA-demethylation activity of AlkB-like proteins evolved to regulate biological RNA methylation, and that the repair of aberrant, chemical methylation is fortuitous.” Ask them how the cell evolved these things, and you’ll probably get a quizzical look, as if “Why are you asking such a dumb question? I don’t know. It just had to. We’re here, aren’t we?”
The Logic of Chance: The Nature and Origin of Biological Evolution, Eugene V. Koonin, page 365:
The RNA World hypothesis faces grave difficulties. First, despite all invested effort, the in vitro evolved ribozymes remain (relatively) poor catalysts for most reactions; the lack of efficient, processive ribozyme polymerases seems particularly troubling, but there is also a serious shortage of other activities, such as those required for the synthesis of nucleotides. Admittedly, it might be unrealistic to expect that experiments on in vitro evolution of ribozymes could easily mimic the actual complexity of the primordial RNA World. Although these experiments harness the power of selection, they are obviously performed on a totally different time scale and under conditions that cannot accurately reproduce the (unknown) conditions at the origin of life
The overall situation in the origin of life field appears rather grim. Even under the (highly nontrivial) assumption that monomers such as NTP are readily available, the problem of the synthesis of sufficiently stable, structurally regular polymers (RNA) is formidable, and the origin of replication and translation from such primordial RNA molecules could be an even harder problem. As emphasized repeatedly in this book, evolution by natural selection
and drift can begin only after replication with sufficient fidelity is established. Even at that stage, the evolution of translation remains highly problematic.

Last edited by Admin on Wed Sep 12, 2018 8:17 am; edited 19 times in total