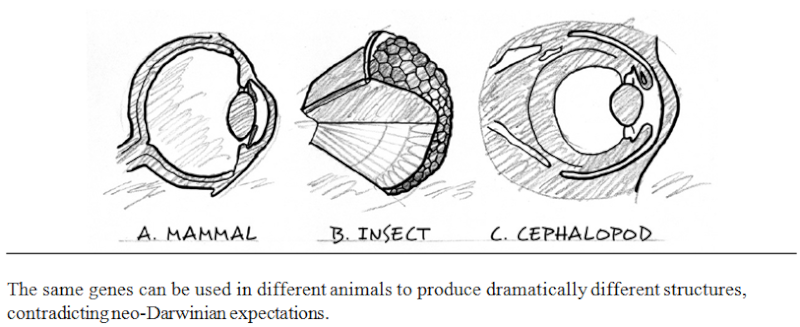
Epigenetics
https://reasonandscience.catsboard.com/t2098-epigenetics
Moshe Szyf: The early life environment and the epigenome 2009 Feb 3
The genome is programmed by the epigenome, which is comprised of chromatin, a covalent modification of DNA by methylation and noncoding RNAs. The epigenome is sculpted during gestation, resulting in the diversity of gene expression programs in the distinct cell types of the organism. Recent data suggest that epigenetic programming of gene expression profiles is sensitive to the early-life environment and that both the chemical and social environment early in life could affect the manner by which the genome is programmed by the epigenome.
https://pubmed.ncbi.nlm.nih.gov/19364482/
María A. Sánchez-Romero: The bacterial epigenome 14 November 2019
In all domains of life, genomes contain epigenetic information superimposed over the nucleotide sequence. Epigenetic signals control DNA–protein interactions and can cause phenotypic change in the absence of mutation. A nearly universal mechanism of epigenetic signalling is DNA methylation. In bacteria, DNA methylation has roles in genome defence, chromosome replication and segregation, nucleoid organization, cell cycle control, DNA repair and regulation of transcription. In many bacterial species, DNA methylation controls reversible switching (phase variation) of gene expression, a phenomenon that generates phenotypic cell variants. The formation of epigenetic lineages enables the adaptation of bacterial populations to harsh or changing environments and modulates the interaction of pathogens with their eukaryotic hosts.
https://www.nature.com/articles/s41579-019-0286-2
Madhumita Bhattacharyya: Origin and Evolution of DNA methyltransferases (DNMT) along the tree of life: A multi-genome survey April 09, 2020.
DNA methylation was found to play important roles in the biology of bacteria: phenomena such as timing of DNA replication, partitioning nascent chromosomes to daughter cells, repair of DNA, and timing of transposition and conjugal transfer of plasmids are sensitive to the methylation states of specific DNA regions.
https://www.biorxiv.org/content/10.1101/2020.04.09.033167v1.full
Thomas Malone: Structure-guided Analysis Reveals Nine Sequence Motifs Conserved among DNA Amino-methyltransferases, and Suggests a Catalytic Mechanism for these Enzymes 1995 Nov 3
DNA Mtases transfer methyl groups from S-adenosyl-L-methionine (AdoMet) to specific positions on bases in double-stranded DNA. The DNA Mtases fall into two major classes, defined by the position methylated. The members of one class methylate a pyrimidine ring carbon yielding C5-methylcytosine (5mC; e.g. HhaI Mtase, M.HhaI). Members of the second class methylate exocyclic amino nitrogens, forming either N6-methyladenine (N6mA; e.g. M.TaqI) or N4-methylcytosine (N4mC; e.g. M.PvuII). Mtases of the two classes were expected to be substantially different from one another, based on the fact that their targets of methyl transfer are very different
https://sci-hub.ee/10.1006/jmbi.1995.0577
Claim: The recent discoveries of epigenetics add to the information at hand. It does not overturn it. There is still population genetics. Mutation, natural selection, genetic drift, non-random mating, and gene flow still happen and still cause population gene pools to change. Allopatric speciation and sympatric speciation still happen due to origins of reproductive-isolating mechanisms. Macroevolution and common descent still exists and still have their supporting evidences. None of that has gone away with the discovery of epigenetic inheritance.
Answer: Epigenetics shift the essential mechanisms that shape cell size and form, body architecture and form, organization and size, to mechanisms beyond Genes. This means as well, in order for form or size either on cell or organ, or organism level to change, epigenetic change has to be taken into consideration. If epigenetic informational codes change, then cells and bodies can and do change as well. This shift is considerable, and essential, and refutes gene-centric views on which the theory of evolution is based.
Following 23 items below do define Cell, organ and organism development, size, type, architecture, and shape:
Genetic:
1.DNA Code sequence
Epigenetic ( beyond or outside the genetic information ) :
1. The 31 Genetic Codes
2. The Adhesion Code
3. The Apoptosis Code
4. The Bioelectric Code
5. The Biophoton Code
6. The Calcium Code
7. The Chaperone Code
8. The Chromatin Code
9. The Circular motif ( ribosome) Code
10. The Coactivator/corepressor/epigenetic Code
11. The Code of human language
12. The Hidden Code within the Genetic Code
13. The DNA methylation Code
14. The Differentiation Code
15. The Domain substrate specificity Code of Nonribosomal peptide synthetases (NRPS)
16. The Error correcting Code
17. The Genomic regulatory Code
18. The Glycomic Code
19. The Histone Code
20. The HOX Code
21. The immune response code, or language
22. The Lamin Code
23. The MeshCODE
24. The Metabolic Code
25. The Myelin Code
26. The Neuronal spike-rate Code
27. The Non-ribosomal Code
28. The Nucleosome Code
29. The Olfactory Code
30. The Operon Code
31. The Phosphorylation Code
32. The Post-translational modification Code for transcription factors
33. The RNA Code
34. The Ribosomal Code
35. The Riboswitch Code
36. The Splicing Codes
37. The Signal transduction Code
38. The Signal Integration Codes
39. The Sugar Code
40. The Synaptic Adhesive Code
41. The Talin Code
42. The Transcription factor Code
43. The Transcriptional cis-regulatory Code
44. The Tubulin Code
45. The Ubiquitin Code
There is just ONE item which is genetic, namely the genetic DNA sequence, which upon population genetics. Mutation, natural selection, genetic drift, non-random mating, and gene flow cause population gene pools to change. Allopatric speciation and sympatric speciation still happen due to origins of reproductive-isolating mechanisms. But these mechanisms on a genetic level have influence in a VERY limited range.
Where Do Complex Organisms Come From?
https://reasonandscience.catsboard.com/t2316-where-do-complex-organisms-come-from
The argument of the genetic piano
1. Dr. Kohzoh Mitsuya [University of Texas Health Science Center] who studies genes says the work of epigenetics “corresponds to a pianist playing a piece of music. Like keys on a piano, DNA is the static blueprint for all the proteins that cells produce.”
2. “Epigenetic information provides additional dynamic or flexible instructions as to how, where and when the blueprint will be used.”
3. After watching the response of mice deficient in the RNA, he said, “It shows how one note is played on the piano. The symphony has only just come into view. We can hear it, but we need to learn how all the parts are being played.”
4. Here the questions are: who’s the pianist and who’s the conductor?
5. The environment cannot be the musician; it is oblivious to the needs of the organism. Heredity cannot be the musician; it has no foresight to read or comprehend a collection of processes organized into a work.
6. Thus, this discovery and explanation of Dr. Mitsuya causes trouble for Darwin while it fits precisely into the intelligent design theory.
7. There must be an origin of the information required to produce function.
8. A classical answer to this by the evolutionists is: “this evolved, that’s why it is there.”
9. Answering this we say: “Science is supposed to seek efficient causes, not just-so stories or appeals to chance based on circular reasoning. For example, in his book The Making of the Fittest, Sean Carroll writes “the degree of similarity in DNA is an index of the [evolutionary] relatedness of species.” [98] This can only make sense if we first assume evolution is true. But Carroll’s book is a defense of evolution, intended to demonstrate that the theory is true without first assuming it is true. He seeks to prove evolution is true, but he begins with evolutionary reasoning and interpretations. That is circular reasoning.”
10. The alternative and only explanation is therefore intelligent design with a known cause sufficient to produce functional information: intelligence. Only intelligence can organize atoms or building blocks into order and activities. There is no other experience of anything else putting things into order and motion.
11. Intelligent design means intelligence of the greatest scientist all men call God.
12. A Creator, most probably, is required, and exists.
Reference:
1. Watanabe, Tomizami, Mitsuya et al, “Role for piRNAs and Noncoding RNA in de Novo DNA Methylation of the Imprinted Mouse Rasgrf1 Locus,” Science, 13 May 2011: Vol. 332 no. 6031 pp. 848-852, DOI: 10.1126/science.1203919.
Humans have only 21,000 genes—the same as a worm—and they are identical in all of the different types of cells. It is not the inherited code of the genes that determines the different cellular functions. Rather, it is way that genes are utilized differently in each type of cell that determines which proteins will produce unique structures. 6
Increasingly, these “epigenetic” mechanisms (that is, mechanisms outside of the simple procedure of assigning an amino acid directly from a code in a gene) are being found to be vastly more complex than ever imagined. This post will describe recent discoveries of dynamic three-dimensional structures in the cell’s nucleus that along with unique localization and packaging of the DNA are vital for every aspect of gene function. Vast complexity of chromatin 3D shapes is another way that DNA is regulated.
Stephen C Meyer , Darwin's doubt pg.218:
Contemporary critics of neo-Darwinism acknowledge, of course, that preexisting forms of life can diversify under the twin influences of natural selection and genetic mutation. Known microevolutionary processes can account for small changes in the coloring of peppered moths, the acquisition of antibiotic resistance in different strains of bacteria, and cyclical variations in the size of Galápagos finch beaks. Nevertheless, many biologists now argue that neo-Darwinian theory does not provide an adequate explanation for the origin of new body plans or events such as the Cambrian explosion. For example, evolutionary biologist Keith Stewart Thomson, formerly of Yale University, has expressed doubt that large-scale morphological changes could accumulate by minor changes at the genetic level. Geneticist George Miklos, of the Australian National University, has argued that neo- Darwinism fails to provide a mechanism that can produce large-scale innovations in form and structure. Biologists Scott Gilbert, John Opitz, and Rudolf Raff have attempted to develop a new theory of evolution to supplement classical neo-Darwinism, which, they argue, cannot adequately explain large-scale macroevolutionary change. As they note:
Starting in the 1970s, many biologists began questioning its neo-Darwinism's adequacy in explaining evolution. Genetics might be adequate for explaining microevolution, but microevolutionary changes in gene frequency were not seen as able to turn a reptile into a mammal or to convert a fish into an amphibian. Microevolution looks at adaptations that concern the survival of the fittest, not the arrival of the fittest. As Goodwin (1995) points out, "the origin of species—Darwin's problem—remains unsolved."
pg. 204
Genes alone do not determine the three-dimensional form and structure of an animal. so-called epigenetic information—information stored in cell structures, but not in DNA sequences—plays a crucial role. The Greek prefix epi means "above" or "beyond," so epigenetics refers to a source of information that lies beyond the genes. "Detailed information at the level of the gene does not serve to explain form." "epigenetic" or "contextual information" plays a crucial role in the formation of animal "body assemblies" during embryological development.
Recent discoveries about the role of epigenetic information in animal development pose a formidable challenge to the standard neo-Darwinian account of the origin of these body plans—perhaps the most formidable of all. "the neo-Darwinian paradigm still represents the central explanatory framework of evolution," it has "no theory of the generative." neo-Darwinism "completely avoids the question of the origination of phenotypic traits and of organismal form." 1
Neo-Darwinism lacks an explanation for the origin of organismal form precisely because it cannot explain the origin of epigenetic information.
FORM AND INFORMATION
Biologists typically define "form" as a distinctive shape and arrangement of body parts. Forms exist in three spatial dimensions and arise in time—in the case of animals during development from embryo to adult. Animal form arises as material constituents are constrained to establish specific arrangements with an identifiable three-dimensional shape , one that we would recognize as the body plan of a particular type of animal. A particular "form," therefore, represents a highly specific arrangement of material components among a much larger set of possible arrangements.
Similarly, animal body plans represent, not only highly improbable, but also highly specific arrangements of matter. Organismal form and function depend upon the precise arrangement of various constituents as they arise during, or contribute to, embryological development. Thus, the specific arrangement of the other building blocks of biological form—cells, clusters of similar cell types, tissues, and organs—also represent a kind of specified or functional information.
Neo-Darwinists have assumed that genes possess all the information necessary to specify the form of an animal. They have also assumed that mutations in genes will suffice to generate the new information necessary to build a new form of animal life. 2
BEYOND GENES
Many biologists no longer believe that DNA directs virtually everything happening within the cell. Developmental biologists, in particular, are now discovering more and more ways that crucial information for building body plans is imparted by the form and structure of embryonic cells, including information from both the unfertilized and fertilized egg. Biologists now refer to these sources of information as "epigenetic." The information needed to code for complex biological systems vastly outstrips the information in DNA. 3
Once proteins are synthesized, they must be arranged into higher-level systems of proteins and structures.
Distinctive cell types are made of, among other things, systems of specialized proteins. Organs are made of specialized arrangements of cell types and tissues. And body plans comprise specific arrangements of specialized organs. Yet the properties of individual proteins do not fully determine the organization of these higher-level structures and patterns. 4
CYTOSKELETAL ARRAYS
Other sources of information must help arrange individual proteins into systems of proteins, systems of proteins into distinctive cell types, cell types into tissues, and different tissues into organs. And different organs and tissues must be arranged to form body plans.
At a construction site, builders will make use of many materials: lumber, wires, nails, drywall, piping, and windows. Yet building materials do not determine the floor plan of the house or the arrangement of houses in a neighborhood. Similarly, electronic circuits are composed of many components, such as resistors, capacitors, and transistors. But such lower-level components do not determine their own arrangement in an integrated circuit
In a similar way, DNA does not by itself direct how individual proteins are assembled into these larger systems or structures—cell types, tissues, organs, and body plans—during animal development.Instead, the three-dimensional structure or spatial architecture of embryonic cells plays important roles in determining body-plan formation during embryogenesis. Developmental biologists have identified several sources of epigenetic information in these cells.
The structure and location of the microtubules in the cytoskeleton influence the patterning and development of embryos. Microtubule "arrays" within embryonic cells help to distribute essential proteins used during development to specific locations in these cells. Once delivered, these proteins perform functions critical to development, but they can only do so if they are delivered to their correct locations with the help of preexisting, precisely structured microtubule or cytoskeletal arrays.
Thus, the precise arrangement of microtubules in the cytoskeleton constitutes a form of critical structural information. These microtubule arrays are made of proteins called tubulin, which are gene products. Nevertheless, like bricks that can be used to assemble many different structures, the tubulin proteins in the cell's microtubules are identical to one another. Thus, neither the tubulin subunits, nor the genes that produce them, account for the differences in the shape of the microtubule arrays that distinguish different kinds of embryos and developmental pathways. Instead, the structure of the microtubule array itself is, once again, determined by the location and arrangement of its subunits, not the properties of the subunits themselves.
The Centrosome
https://www.youtube.com/watch?v=5rqbmLiSkpk
Another cell structure influences the arrangement of the microtubule arrays and thus the precise structures they form and the functions they perform. In an animal cell, that structure is called the centrosome (literally, "central body"), a microscopic organelle that sits next to the nucleus between cell divisions in an undividing cell. Emanating from the centrosome is the microtubule array that gives a cell its three-dimensional shape and provides internal tracks for the directed transport of organelles and essential molecules to and from the nucleus. During cell division the centrosome duplicates itself. The two centrosomes form the poles of the cell-division apparatus, and each daughter cell inherits one of the centrosomes; yet the centrosome contains no DNA. Though centrosomes are made of proteins—gene products—the centrosome structure is not determined by genes alone.
pg 213:
NEO-DARWINISM AND THE CHALLENGE OF EPIGENETIC INFORMATION
These different sources of epigenetic information in embryonic cells pose an enormous challenge to the sufficiency of the neo-Darwinian mechanism. According to neo-Darwinism, new information, form, and structure arise from natural selection acting on random mutations arising at a very low level within the biological hierarchy—within the genetic text. Yet both body-plan formation during embryological development and major morphological innovation during the history of life depend upon a specificity of arrangement at a much higher level of the organizational hierarchy, a level that DNA alone does not determine. If DNA isn't wholly responsible for the way an embryo develops— for body-plan morphogenesis—then DNA sequences can mutate indefinitely and still not produce a new body plan, regardless of the amount of time and the number of mutational trials available to the evolutionary process. Genetic mutations are simply the wrong tool for the job at hand.
Even in a best-case scenario—one that ignores the immense improbability of generating new genes by mutation and selection—mutations in DNA sequence would merely produce new genetic information. But building a new body plan requires more than just genetic information. It requires both genetic and epigenetic information—information by definition that is not stored in DNA and thus cannot be generated by mutations to the DNA. It follows that the mechanism of natural selection acting on random mutations in DNA cannot by itself generate novel body plans, such as those that first arose in the Cambrian explosion.
In at least the case of the sugar molecules on the cell surface, gene products play no direct role. Genetic information produces proteins and RNA molecules, not sugars and carbohydrates. Of course, important glycoproteins and glycolipids (sugar-protein and sugar-fat composite molecules) are modified as the result of biosynthetic pathways involving networks of proteins. Nevertheless, the genetic information that generates the proteins in these pathways only determines the function and structure of the individual proteins; it does not specify the coordinated interaction between the proteins in the pathways that result in the modification of sugars
Some biologists have noted that so-called helper proteins—which are gene products—called "microtubule associated proteins" (MAPs) help to assemble the tubulin subunits in the microtubule arrays. Yet MAPs, and indeed many other necessary proteins, are only part of the story. The locations of specified target sites on the interior of the cell membrane also help to determine the shape of the cytoskeleton. And, as noted, the gene products out of which these targets are made do not determine the location of these targets. Similarly, the position and structure of the centrosome—the microtubule- organizing center—also influences the structure of the cytoskeleton. Although centrosomes are made of proteins, the proteins that form these structures do not entirely determine their location and form.
As Mark McNiven, a molecular biologist at the Mayo Clinic, and cell biologist Keith Porter, formerly of the University of Colorado, have shown, centrosome structure and membrane patterns as a whole convey three-dimensional structural information that helps determine the structure of the cytoskeleton and the location of its subunits.
In each new generation, the form and structure of the cell arises as the result of both gene products and the preexisting three-dimensional structure and organization inherent in cells, cell membranes, and cyto-skeletons. Many cellular structures are built from proteins, but proteins find their way to correct locations in part because of preexisting three-dimensional patterns and organization inherent in cellular structures. Neither structural proteins nor the genes that code for them can alone determine the three-dimensional shape and structure of the entities they build. Gene products provide necessary, but not sufficient, conditions for the development of three-dimensional structure within cells, organs, and body plans. 5 If this is so, then natural selection acting on genetic variation and mutations alone cannot produce the new forms that arise in the history of life.
The structures in which epigenetic information inheres—cytoskeletal arrays and membrane patterns, for example—are much larger than individual nucleotide bases or even stretches of DNA. For this reason, these structures are not vulnerable to alteration by many of the typical sources of mutation that affect genes such as radiation and chemical agent. To the extent that cell structures can be altered, these alterations are overwhelmingly likely to have harmful or catastrophic consequences.
1) Müller and Newman, "Origination of Organismal Form," Or as Müller also explains, the
question of how "individualized constructional elements" are organized during "the evolution of
organismal form" is "not satisfactorily answered by current evolutionary theories"; Müller, "Homology," 57-58
2) Levinton, Genetics, Paleontology, and Macroevolution, 485
3) Goodwin, "What Are the Causes of Morphogenesis?"; Nijhout, "Metaphors and the Role of Genes in Development"; Sapp, Beyond the Gene; Müller and Newman, "Origination of Organismal Form"; Brenner, "The Genetics of Behaviour"; Harold, The Way of the Cell.
4) Harold, The Way of the Cell, 125.
5) Harold, "From Morphogenes to Morphogenesis," 2767
http://www.microbiologyresearch.org/docserver/fulltext/micro/141/11/mic-141-11-2765.pdf expires=1475512019&id=id&accname=guest&checksum=82B036E71F67093D4C4B4F076E6D8114
Harold, “From Morphogenes to Morphogenesis,” 2774; Moss, What Genes Can’t Do. Of course, many proteins bind chemically with each other to form complexes and structures within cells. Nevertheless, these “self-organizational” properties do not fully account for higher levels of organization in cells, organs, or body plans. Or, as Moss has explained “Neither DNA nor any other aperiodic crystal constitutes a unique repository of heritable stability in the cell; in addition, the chemistry of the solid state does not constitute either a unique or even an ontologically or causally privileged basis for explaining the existence and continuity of order in the living world . . .” Moss, What Genes Can’t Do, 76.
Morphogenesis cannot be orchestrated by the genome, but makes manifest a higher level of order, corresponding to the cellular scale of size and organization. If a single phrase can stand for the whole riddle, it may be ‘ cell polarity ’, The term refers to the visible directionality
6) http://jonlieffmd.com/blog/vast-complexity-of-chromatin-3d-shapes
https://reasonandscience.catsboard.com/t2098-epigenetics
Moshe Szyf: The early life environment and the epigenome 2009 Feb 3
The genome is programmed by the epigenome, which is comprised of chromatin, a covalent modification of DNA by methylation and noncoding RNAs. The epigenome is sculpted during gestation, resulting in the diversity of gene expression programs in the distinct cell types of the organism. Recent data suggest that epigenetic programming of gene expression profiles is sensitive to the early-life environment and that both the chemical and social environment early in life could affect the manner by which the genome is programmed by the epigenome.
https://pubmed.ncbi.nlm.nih.gov/19364482/
María A. Sánchez-Romero: The bacterial epigenome 14 November 2019
In all domains of life, genomes contain epigenetic information superimposed over the nucleotide sequence. Epigenetic signals control DNA–protein interactions and can cause phenotypic change in the absence of mutation. A nearly universal mechanism of epigenetic signalling is DNA methylation. In bacteria, DNA methylation has roles in genome defence, chromosome replication and segregation, nucleoid organization, cell cycle control, DNA repair and regulation of transcription. In many bacterial species, DNA methylation controls reversible switching (phase variation) of gene expression, a phenomenon that generates phenotypic cell variants. The formation of epigenetic lineages enables the adaptation of bacterial populations to harsh or changing environments and modulates the interaction of pathogens with their eukaryotic hosts.
https://www.nature.com/articles/s41579-019-0286-2
Madhumita Bhattacharyya: Origin and Evolution of DNA methyltransferases (DNMT) along the tree of life: A multi-genome survey April 09, 2020.
DNA methylation was found to play important roles in the biology of bacteria: phenomena such as timing of DNA replication, partitioning nascent chromosomes to daughter cells, repair of DNA, and timing of transposition and conjugal transfer of plasmids are sensitive to the methylation states of specific DNA regions.
https://www.biorxiv.org/content/10.1101/2020.04.09.033167v1.full
Thomas Malone: Structure-guided Analysis Reveals Nine Sequence Motifs Conserved among DNA Amino-methyltransferases, and Suggests a Catalytic Mechanism for these Enzymes 1995 Nov 3
DNA Mtases transfer methyl groups from S-adenosyl-L-methionine (AdoMet) to specific positions on bases in double-stranded DNA. The DNA Mtases fall into two major classes, defined by the position methylated. The members of one class methylate a pyrimidine ring carbon yielding C5-methylcytosine (5mC; e.g. HhaI Mtase, M.HhaI). Members of the second class methylate exocyclic amino nitrogens, forming either N6-methyladenine (N6mA; e.g. M.TaqI) or N4-methylcytosine (N4mC; e.g. M.PvuII). Mtases of the two classes were expected to be substantially different from one another, based on the fact that their targets of methyl transfer are very different
https://sci-hub.ee/10.1006/jmbi.1995.0577
Claim: The recent discoveries of epigenetics add to the information at hand. It does not overturn it. There is still population genetics. Mutation, natural selection, genetic drift, non-random mating, and gene flow still happen and still cause population gene pools to change. Allopatric speciation and sympatric speciation still happen due to origins of reproductive-isolating mechanisms. Macroevolution and common descent still exists and still have their supporting evidences. None of that has gone away with the discovery of epigenetic inheritance.
Answer: Epigenetics shift the essential mechanisms that shape cell size and form, body architecture and form, organization and size, to mechanisms beyond Genes. This means as well, in order for form or size either on cell or organ, or organism level to change, epigenetic change has to be taken into consideration. If epigenetic informational codes change, then cells and bodies can and do change as well. This shift is considerable, and essential, and refutes gene-centric views on which the theory of evolution is based.
Following 23 items below do define Cell, organ and organism development, size, type, architecture, and shape:
Genetic:
1.DNA Code sequence
Epigenetic ( beyond or outside the genetic information ) :
1. The 31 Genetic Codes
2. The Adhesion Code
3. The Apoptosis Code
4. The Bioelectric Code
5. The Biophoton Code
6. The Calcium Code
7. The Chaperone Code
8. The Chromatin Code
9. The Circular motif ( ribosome) Code
10. The Coactivator/corepressor/epigenetic Code
11. The Code of human language
12. The Hidden Code within the Genetic Code
13. The DNA methylation Code
14. The Differentiation Code
15. The Domain substrate specificity Code of Nonribosomal peptide synthetases (NRPS)
16. The Error correcting Code
17. The Genomic regulatory Code
18. The Glycomic Code
19. The Histone Code
20. The HOX Code
21. The immune response code, or language
22. The Lamin Code
23. The MeshCODE
24. The Metabolic Code
25. The Myelin Code
26. The Neuronal spike-rate Code
27. The Non-ribosomal Code
28. The Nucleosome Code
29. The Olfactory Code
30. The Operon Code
31. The Phosphorylation Code
32. The Post-translational modification Code for transcription factors
33. The RNA Code
34. The Ribosomal Code
35. The Riboswitch Code
36. The Splicing Codes
37. The Signal transduction Code
38. The Signal Integration Codes
39. The Sugar Code
40. The Synaptic Adhesive Code
41. The Talin Code
42. The Transcription factor Code
43. The Transcriptional cis-regulatory Code
44. The Tubulin Code
45. The Ubiquitin Code
There is just ONE item which is genetic, namely the genetic DNA sequence, which upon population genetics. Mutation, natural selection, genetic drift, non-random mating, and gene flow cause population gene pools to change. Allopatric speciation and sympatric speciation still happen due to origins of reproductive-isolating mechanisms. But these mechanisms on a genetic level have influence in a VERY limited range.
Where Do Complex Organisms Come From?
https://reasonandscience.catsboard.com/t2316-where-do-complex-organisms-come-from
The argument of the genetic piano
1. Dr. Kohzoh Mitsuya [University of Texas Health Science Center] who studies genes says the work of epigenetics “corresponds to a pianist playing a piece of music. Like keys on a piano, DNA is the static blueprint for all the proteins that cells produce.”
2. “Epigenetic information provides additional dynamic or flexible instructions as to how, where and when the blueprint will be used.”
3. After watching the response of mice deficient in the RNA, he said, “It shows how one note is played on the piano. The symphony has only just come into view. We can hear it, but we need to learn how all the parts are being played.”
4. Here the questions are: who’s the pianist and who’s the conductor?
5. The environment cannot be the musician; it is oblivious to the needs of the organism. Heredity cannot be the musician; it has no foresight to read or comprehend a collection of processes organized into a work.
6. Thus, this discovery and explanation of Dr. Mitsuya causes trouble for Darwin while it fits precisely into the intelligent design theory.
7. There must be an origin of the information required to produce function.
8. A classical answer to this by the evolutionists is: “this evolved, that’s why it is there.”
9. Answering this we say: “Science is supposed to seek efficient causes, not just-so stories or appeals to chance based on circular reasoning. For example, in his book The Making of the Fittest, Sean Carroll writes “the degree of similarity in DNA is an index of the [evolutionary] relatedness of species.” [98] This can only make sense if we first assume evolution is true. But Carroll’s book is a defense of evolution, intended to demonstrate that the theory is true without first assuming it is true. He seeks to prove evolution is true, but he begins with evolutionary reasoning and interpretations. That is circular reasoning.”
10. The alternative and only explanation is therefore intelligent design with a known cause sufficient to produce functional information: intelligence. Only intelligence can organize atoms or building blocks into order and activities. There is no other experience of anything else putting things into order and motion.
11. Intelligent design means intelligence of the greatest scientist all men call God.
12. A Creator, most probably, is required, and exists.
Reference:
1. Watanabe, Tomizami, Mitsuya et al, “Role for piRNAs and Noncoding RNA in de Novo DNA Methylation of the Imprinted Mouse Rasgrf1 Locus,” Science, 13 May 2011: Vol. 332 no. 6031 pp. 848-852, DOI: 10.1126/science.1203919.
Humans have only 21,000 genes—the same as a worm—and they are identical in all of the different types of cells. It is not the inherited code of the genes that determines the different cellular functions. Rather, it is way that genes are utilized differently in each type of cell that determines which proteins will produce unique structures. 6
Increasingly, these “epigenetic” mechanisms (that is, mechanisms outside of the simple procedure of assigning an amino acid directly from a code in a gene) are being found to be vastly more complex than ever imagined. This post will describe recent discoveries of dynamic three-dimensional structures in the cell’s nucleus that along with unique localization and packaging of the DNA are vital for every aspect of gene function. Vast complexity of chromatin 3D shapes is another way that DNA is regulated.
Stephen C Meyer , Darwin's doubt pg.218:
Contemporary critics of neo-Darwinism acknowledge, of course, that preexisting forms of life can diversify under the twin influences of natural selection and genetic mutation. Known microevolutionary processes can account for small changes in the coloring of peppered moths, the acquisition of antibiotic resistance in different strains of bacteria, and cyclical variations in the size of Galápagos finch beaks. Nevertheless, many biologists now argue that neo-Darwinian theory does not provide an adequate explanation for the origin of new body plans or events such as the Cambrian explosion. For example, evolutionary biologist Keith Stewart Thomson, formerly of Yale University, has expressed doubt that large-scale morphological changes could accumulate by minor changes at the genetic level. Geneticist George Miklos, of the Australian National University, has argued that neo- Darwinism fails to provide a mechanism that can produce large-scale innovations in form and structure. Biologists Scott Gilbert, John Opitz, and Rudolf Raff have attempted to develop a new theory of evolution to supplement classical neo-Darwinism, which, they argue, cannot adequately explain large-scale macroevolutionary change. As they note:
Starting in the 1970s, many biologists began questioning its neo-Darwinism's adequacy in explaining evolution. Genetics might be adequate for explaining microevolution, but microevolutionary changes in gene frequency were not seen as able to turn a reptile into a mammal or to convert a fish into an amphibian. Microevolution looks at adaptations that concern the survival of the fittest, not the arrival of the fittest. As Goodwin (1995) points out, "the origin of species—Darwin's problem—remains unsolved."
pg. 204
Genes alone do not determine the three-dimensional form and structure of an animal. so-called epigenetic information—information stored in cell structures, but not in DNA sequences—plays a crucial role. The Greek prefix epi means "above" or "beyond," so epigenetics refers to a source of information that lies beyond the genes. "Detailed information at the level of the gene does not serve to explain form." "epigenetic" or "contextual information" plays a crucial role in the formation of animal "body assemblies" during embryological development.
Recent discoveries about the role of epigenetic information in animal development pose a formidable challenge to the standard neo-Darwinian account of the origin of these body plans—perhaps the most formidable of all. "the neo-Darwinian paradigm still represents the central explanatory framework of evolution," it has "no theory of the generative." neo-Darwinism "completely avoids the question of the origination of phenotypic traits and of organismal form." 1
Neo-Darwinism lacks an explanation for the origin of organismal form precisely because it cannot explain the origin of epigenetic information.
FORM AND INFORMATION
Biologists typically define "form" as a distinctive shape and arrangement of body parts. Forms exist in three spatial dimensions and arise in time—in the case of animals during development from embryo to adult. Animal form arises as material constituents are constrained to establish specific arrangements with an identifiable three-dimensional shape , one that we would recognize as the body plan of a particular type of animal. A particular "form," therefore, represents a highly specific arrangement of material components among a much larger set of possible arrangements.
Similarly, animal body plans represent, not only highly improbable, but also highly specific arrangements of matter. Organismal form and function depend upon the precise arrangement of various constituents as they arise during, or contribute to, embryological development. Thus, the specific arrangement of the other building blocks of biological form—cells, clusters of similar cell types, tissues, and organs—also represent a kind of specified or functional information.
Neo-Darwinists have assumed that genes possess all the information necessary to specify the form of an animal. They have also assumed that mutations in genes will suffice to generate the new information necessary to build a new form of animal life. 2
BEYOND GENES
Many biologists no longer believe that DNA directs virtually everything happening within the cell. Developmental biologists, in particular, are now discovering more and more ways that crucial information for building body plans is imparted by the form and structure of embryonic cells, including information from both the unfertilized and fertilized egg. Biologists now refer to these sources of information as "epigenetic." The information needed to code for complex biological systems vastly outstrips the information in DNA. 3
Once proteins are synthesized, they must be arranged into higher-level systems of proteins and structures.
Distinctive cell types are made of, among other things, systems of specialized proteins. Organs are made of specialized arrangements of cell types and tissues. And body plans comprise specific arrangements of specialized organs. Yet the properties of individual proteins do not fully determine the organization of these higher-level structures and patterns. 4
CYTOSKELETAL ARRAYS
Other sources of information must help arrange individual proteins into systems of proteins, systems of proteins into distinctive cell types, cell types into tissues, and different tissues into organs. And different organs and tissues must be arranged to form body plans.
At a construction site, builders will make use of many materials: lumber, wires, nails, drywall, piping, and windows. Yet building materials do not determine the floor plan of the house or the arrangement of houses in a neighborhood. Similarly, electronic circuits are composed of many components, such as resistors, capacitors, and transistors. But such lower-level components do not determine their own arrangement in an integrated circuit
In a similar way, DNA does not by itself direct how individual proteins are assembled into these larger systems or structures—cell types, tissues, organs, and body plans—during animal development.Instead, the three-dimensional structure or spatial architecture of embryonic cells plays important roles in determining body-plan formation during embryogenesis. Developmental biologists have identified several sources of epigenetic information in these cells.
The structure and location of the microtubules in the cytoskeleton influence the patterning and development of embryos. Microtubule "arrays" within embryonic cells help to distribute essential proteins used during development to specific locations in these cells. Once delivered, these proteins perform functions critical to development, but they can only do so if they are delivered to their correct locations with the help of preexisting, precisely structured microtubule or cytoskeletal arrays.
Thus, the precise arrangement of microtubules in the cytoskeleton constitutes a form of critical structural information. These microtubule arrays are made of proteins called tubulin, which are gene products. Nevertheless, like bricks that can be used to assemble many different structures, the tubulin proteins in the cell's microtubules are identical to one another. Thus, neither the tubulin subunits, nor the genes that produce them, account for the differences in the shape of the microtubule arrays that distinguish different kinds of embryos and developmental pathways. Instead, the structure of the microtubule array itself is, once again, determined by the location and arrangement of its subunits, not the properties of the subunits themselves.
The Centrosome
https://www.youtube.com/watch?v=5rqbmLiSkpk
Another cell structure influences the arrangement of the microtubule arrays and thus the precise structures they form and the functions they perform. In an animal cell, that structure is called the centrosome (literally, "central body"), a microscopic organelle that sits next to the nucleus between cell divisions in an undividing cell. Emanating from the centrosome is the microtubule array that gives a cell its three-dimensional shape and provides internal tracks for the directed transport of organelles and essential molecules to and from the nucleus. During cell division the centrosome duplicates itself. The two centrosomes form the poles of the cell-division apparatus, and each daughter cell inherits one of the centrosomes; yet the centrosome contains no DNA. Though centrosomes are made of proteins—gene products—the centrosome structure is not determined by genes alone.
pg 213:
NEO-DARWINISM AND THE CHALLENGE OF EPIGENETIC INFORMATION
These different sources of epigenetic information in embryonic cells pose an enormous challenge to the sufficiency of the neo-Darwinian mechanism. According to neo-Darwinism, new information, form, and structure arise from natural selection acting on random mutations arising at a very low level within the biological hierarchy—within the genetic text. Yet both body-plan formation during embryological development and major morphological innovation during the history of life depend upon a specificity of arrangement at a much higher level of the organizational hierarchy, a level that DNA alone does not determine. If DNA isn't wholly responsible for the way an embryo develops— for body-plan morphogenesis—then DNA sequences can mutate indefinitely and still not produce a new body plan, regardless of the amount of time and the number of mutational trials available to the evolutionary process. Genetic mutations are simply the wrong tool for the job at hand.
Even in a best-case scenario—one that ignores the immense improbability of generating new genes by mutation and selection—mutations in DNA sequence would merely produce new genetic information. But building a new body plan requires more than just genetic information. It requires both genetic and epigenetic information—information by definition that is not stored in DNA and thus cannot be generated by mutations to the DNA. It follows that the mechanism of natural selection acting on random mutations in DNA cannot by itself generate novel body plans, such as those that first arose in the Cambrian explosion.
In at least the case of the sugar molecules on the cell surface, gene products play no direct role. Genetic information produces proteins and RNA molecules, not sugars and carbohydrates. Of course, important glycoproteins and glycolipids (sugar-protein and sugar-fat composite molecules) are modified as the result of biosynthetic pathways involving networks of proteins. Nevertheless, the genetic information that generates the proteins in these pathways only determines the function and structure of the individual proteins; it does not specify the coordinated interaction between the proteins in the pathways that result in the modification of sugars
Some biologists have noted that so-called helper proteins—which are gene products—called "microtubule associated proteins" (MAPs) help to assemble the tubulin subunits in the microtubule arrays. Yet MAPs, and indeed many other necessary proteins, are only part of the story. The locations of specified target sites on the interior of the cell membrane also help to determine the shape of the cytoskeleton. And, as noted, the gene products out of which these targets are made do not determine the location of these targets. Similarly, the position and structure of the centrosome—the microtubule- organizing center—also influences the structure of the cytoskeleton. Although centrosomes are made of proteins, the proteins that form these structures do not entirely determine their location and form.
As Mark McNiven, a molecular biologist at the Mayo Clinic, and cell biologist Keith Porter, formerly of the University of Colorado, have shown, centrosome structure and membrane patterns as a whole convey three-dimensional structural information that helps determine the structure of the cytoskeleton and the location of its subunits.
In each new generation, the form and structure of the cell arises as the result of both gene products and the preexisting three-dimensional structure and organization inherent in cells, cell membranes, and cyto-skeletons. Many cellular structures are built from proteins, but proteins find their way to correct locations in part because of preexisting three-dimensional patterns and organization inherent in cellular structures. Neither structural proteins nor the genes that code for them can alone determine the three-dimensional shape and structure of the entities they build. Gene products provide necessary, but not sufficient, conditions for the development of three-dimensional structure within cells, organs, and body plans. 5 If this is so, then natural selection acting on genetic variation and mutations alone cannot produce the new forms that arise in the history of life.
The structures in which epigenetic information inheres—cytoskeletal arrays and membrane patterns, for example—are much larger than individual nucleotide bases or even stretches of DNA. For this reason, these structures are not vulnerable to alteration by many of the typical sources of mutation that affect genes such as radiation and chemical agent. To the extent that cell structures can be altered, these alterations are overwhelmingly likely to have harmful or catastrophic consequences.
1) Müller and Newman, "Origination of Organismal Form," Or as Müller also explains, the
question of how "individualized constructional elements" are organized during "the evolution of
organismal form" is "not satisfactorily answered by current evolutionary theories"; Müller, "Homology," 57-58
2) Levinton, Genetics, Paleontology, and Macroevolution, 485
3) Goodwin, "What Are the Causes of Morphogenesis?"; Nijhout, "Metaphors and the Role of Genes in Development"; Sapp, Beyond the Gene; Müller and Newman, "Origination of Organismal Form"; Brenner, "The Genetics of Behaviour"; Harold, The Way of the Cell.
4) Harold, The Way of the Cell, 125.
5) Harold, "From Morphogenes to Morphogenesis," 2767
http://www.microbiologyresearch.org/docserver/fulltext/micro/141/11/mic-141-11-2765.pdf expires=1475512019&id=id&accname=guest&checksum=82B036E71F67093D4C4B4F076E6D8114
Harold, “From Morphogenes to Morphogenesis,” 2774; Moss, What Genes Can’t Do. Of course, many proteins bind chemically with each other to form complexes and structures within cells. Nevertheless, these “self-organizational” properties do not fully account for higher levels of organization in cells, organs, or body plans. Or, as Moss has explained “Neither DNA nor any other aperiodic crystal constitutes a unique repository of heritable stability in the cell; in addition, the chemistry of the solid state does not constitute either a unique or even an ontologically or causally privileged basis for explaining the existence and continuity of order in the living world . . .” Moss, What Genes Can’t Do, 76.
Morphogenesis cannot be orchestrated by the genome, but makes manifest a higher level of order, corresponding to the cellular scale of size and organization. If a single phrase can stand for the whole riddle, it may be ‘ cell polarity ’, The term refers to the visible directionality
6) http://jonlieffmd.com/blog/vast-complexity-of-chromatin-3d-shapes
Last edited by Otangelo on Sat Jun 11, 2022 8:46 am; edited 15 times in total