


A dinoflagellate protist which has eyes like in vertebrates, and ballistic multi-barrel guns for taking out prey. By design, or evolution?
Two weird dinoflagellate protists have ballistic "nematocyst" organelles. The weaponry is complicated, savage, and unlike what’s seen in animals. These bugs are basically the battle tanks of the microbial world, with armor-piercing, multi-barrel Gatling guns for taking out prey. So cool! 1
Dinoflagellates called Nematodinium and Polykrikos are microscopic plankton, the kind of flotsam that whales gulp up by the ton. But these eukaryotic dinoflagellates, a type of protist, have their own drama-filled lives. They are hunters that eat other dinoflagellates, which themselves are bristling with armor, microscopic munitions and even chemical weapons. The tiny weapons of Nematodinium and related dinoflagellates are a unique invention: Though the weapons look a lot like the stingers of jellyfish, the structures are different. 8
Dinoflagellates are the coolest cells. Warnowiaceae dinoflagellates which also have nematocysts throwing out prey-capturing filaments are among the most complex single-celled organisms we know of. 4
Nematodinium's weird clusters of weaponry are called nematocysts. Gavelis and his colleagues used scanning electron microscopy (SEM) to image these nematocysts as well as the nematocysts of another well-armed dinoflagellate, Polykrikos kofoidii. They also captured the firing of P. kofoidii's weaponry in high-speed video for the first time.
What they saw were some intricately complex structures: Nematodinium has nematocysts that are clustered in rosette shapes and look like the multibarrel configuration of a Gatling gun.
Nobody has ever seen these creatures hunt, because Nematodinium is super-rare. "Unlike a Gatling gun, it probably shoots all of those capsules off at the same time."
Polykrikos kofoidii is a dinoflagellate that eats other dinoflagellates, capturing them with harpoon-like weapons that pierce the prey and drag them in to be consumed. This is a weird mechanism. This dinoflagellate sports capsules on its surface, each of which is topped by a finger-like projection called a taeniocyst. When it comes into contact with prey, the taeniocyst explodes, perhaps shooting out an adhesive similar to Spider-Man's webbing, Gavelis said. The eruption of the taeniocyst, in turn, triggers the capsule, or nematocyst, to shoot out a coiled tube tipped with a pointy, dagger-like projection called a stylet. The stylet pierces its way out of the capsule and penetrates the prey. The coil dissolves, but the stylet is still attached to the predator dinoflagellate by a towline. It uses that towline, basically like someone pulling a harpooned fish to its demise. Then, it's snack time.
Stinging sea animals, such as jellyfish and other cnidarians, have nematocysts, too. For that reason, there have long been theories that perhaps the dinoflagellates and the cnidarians evolved from some long-ago common ancestor with some pretty impressive defenses. Comparisons of the proteins found in cnidarians with those found in dinoflagellates indicates that there are no relationships.
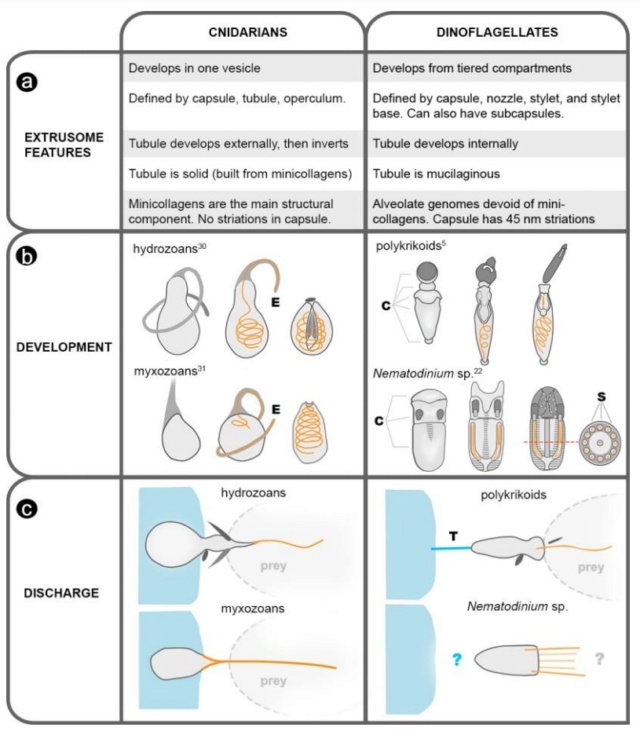
A synthesis of fundamental differences between the nematocysts in cnidarians and dinoflagellates.
(a) A list of extrusomal features (with references given in 5, 22, 30, 31). (b) Illustrations of nematocyst development; tubules are orange, the dotted line in Nematodinium represents the plane of the cross section, on right. E = everting tubule. C = tiered compartments. S = subcapsules. (c) Illustrations of nematocysts firing. All illustrations are inferred from live observation except for that of Nematodinium, in which discharge has been only partially observed, in vitro. T = tow filament.
The genes that build nematocysts in dinoflagellates are entirely different from those that build them in cnidarians.
Biologists sometimes use the phrase "arms race" to describe a tug-of-war, but it's rarely this literal. Microbes called dinoflagellates have intricate weapons—including a microscopic version of a Gatling gun—to harpoon their dinners. Scientists have known about these harpoons for decades, and some have guessed that the weapons stem from the same source as the ones wielded by jellyfish and other cnidarians. An analysis of the genes and proteins involved with weapon construction, however, shows that dinoflagellates and cnidarians use different proteins to manufacture their weapons—meaning they arrived at similar solutions through separate paths. 2
Microbial arms race: Ballistic “nematocysts” in dinoflagellates represent a new extreme in organelle complexity
https://advances.sciencemag.org/content/3/3/e1602552
We examine the origin of harpoon-like secretory organelles (nematocysts) in dinoflagellate protists. These ballistic organelles have been hypothesized to be homologous to similarly complex structures in animals (cnidarians); but we show, using structural, functional, and phylogenomic data, that nematocystsevolved independently in both have different lineages. We also recorded the first high-resolution videos of nematocyst discharge in dinoflagellates. Unexpectedly, our data suggest that different types of dinoflagellate nematocysts use two fundamentally different types of ballistic mechanisms: one type relies on a single pressurized capsule for propulsion, whereas the other type launches 11 to 15 projectiles from an arrangement similar to a Gatling gun. Despite their radical structural differences, these nematocysts share a single origin within dinoflagellates and both potentially use a contraction-based mechanism to generate ballistic force. The diversity of traits in dinoflagellate nematocysts demonstrates a stepwise route by which simple secretory structures diversified to yield elaborate subcellular weaponry.
My comment: Why and how does that demonstrate a stepwise route ? It actually demonstrates the contrary. Namely that a stepwise route is not feasible because the structures are radically different.

Model of nematocyst homology and evolution in dinoflagellates.
(a) Illustration of homologous components in the known nematocysts of dinoflagellates as inferred from single-cell focused ion beam scanning electron microscopy (FIB-SEM) and serial TEM sections (15, 22, 30, 43, 44). Colors indicate homology of specific components of nematocysts (red = nozzle, blue = capsule, black = stylet, yellow = stylet base, purple = tubule, grey = uncertain), and horizontal grey lines indicate homology between the major membrane-bound compartments.
(b) Illustration of nematocyst traits mapped within a molecular phylogenetic context (based on fig. S6) showing the dinoflagellates with complex extrusomes. The number of major nematocyst vesicles/compartments is inferred to have increased over time along the lineage leading to polykrikoids (e.g., Polykrikos) and warnowiids (e.g., Nematodinium); the number of radially arranged subcompartments within nematocysts is inferred to have increased over time along the lineage leading to warnowiids (e.g., Nematodinium sp.).
(B) SEM micrograph of an isolated taeniocyst that has discharged its amorphous contents.
(C) SEM micrograph of an isolated nematocyst that has become arrested very early in discharge; arrowhead, operculum.
(D) FIB-SEM section of taeniocyst (the white thing at the top) and nematocyst enclosed by a membranous chute (arrowhead) and
(E) maximum intensity projection of the same region seen slightly from above and below
(F). (G) Virtual dissection of the nematocyst-taeniocyst complex. Brackets indicate the membrane-bound compartments in which those components are grouped during early development. Later in development, compartments fuse to form the chute.
The capsule or nematocyst
1. Taeniocyst triggers the capsule, or nematocyst, to shoot out the coiled tube when it comes into contact with prey and explodes
2. a "linker" organelle. Use not known
3. a Nozzle designed to control the direction or characteristics of the tube flow as it exits the enclosed chamber
4. a base is where the tubule uncoils and is projected through
5. the stylet is a pointy dagger-like projection which pierces its way out of the capsule and penetrates the prey
6. the coiled Tube is tipped with a pointy, dagger-like projection called a stylet.
7. the capsule is the container of the tubes
8. a towline is used to pull the harpooned prey to its demise.
9. The tow filament is on the proximal end of the nematocyst and likely originates from the posterior vesicle.
This is an integrated interdependent system, where all parts must be in place in order to reach the higher end, that is to catch prey.
This dinoflagellate sports capsules on its surface, each of which is topped by a finger-like projection called a taeniocyst. When it comes into contact with prey, the taeniocyst explodes, perhaps shooting out an adhesive similar to Spider-Man's webbing. The eruption of the taeniocyst, in turn, triggers the capsule, or nematocyst, to shoot out a coiled tube tipped with a pointy, dagger-like projection called a stylet. The stylet pierces its way out of the capsule and penetrates the prey. The coil dissolves, but the stylet is still attached to the predator dinoflagellate by a towline. It uses that towline, basically like someone pulling a harpooned fish to its demise. Then, it's snack time.
The taeniocyst—that emerges from a finger-like projection near the top of each Polykrikos cell. Although the taeniocyst makes first contact with the prey, its functions are unclear. We provide evidence that taeniocysts are also ballistic structures. On five occasions, we observed that taeniocysts violently discharge when isolated from the cell by launching their contents through an apical channel (Fig. below).
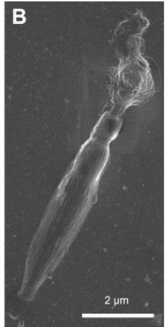
The taeniocyst and nematocyst seem to work in tandem, with
(i) the taeniocyst intially adhering to the prey, followed by
(ii) discharge of the nematocyst, which punctures the prey, and last
(iii), the prey is retrieved using a tow filament.
Each mature nematocyst-taeniocyst complex resides within a single membrane-bound compartment (for example, the “chute”)
A novel “linker” organelle. This cylindrical structure connected the taeniocyst and the nematocyst further indicating that these two extrusomes types are deployed in succession, in a coordinated ballistic response. No comparable arrangement is found in cnidarians.
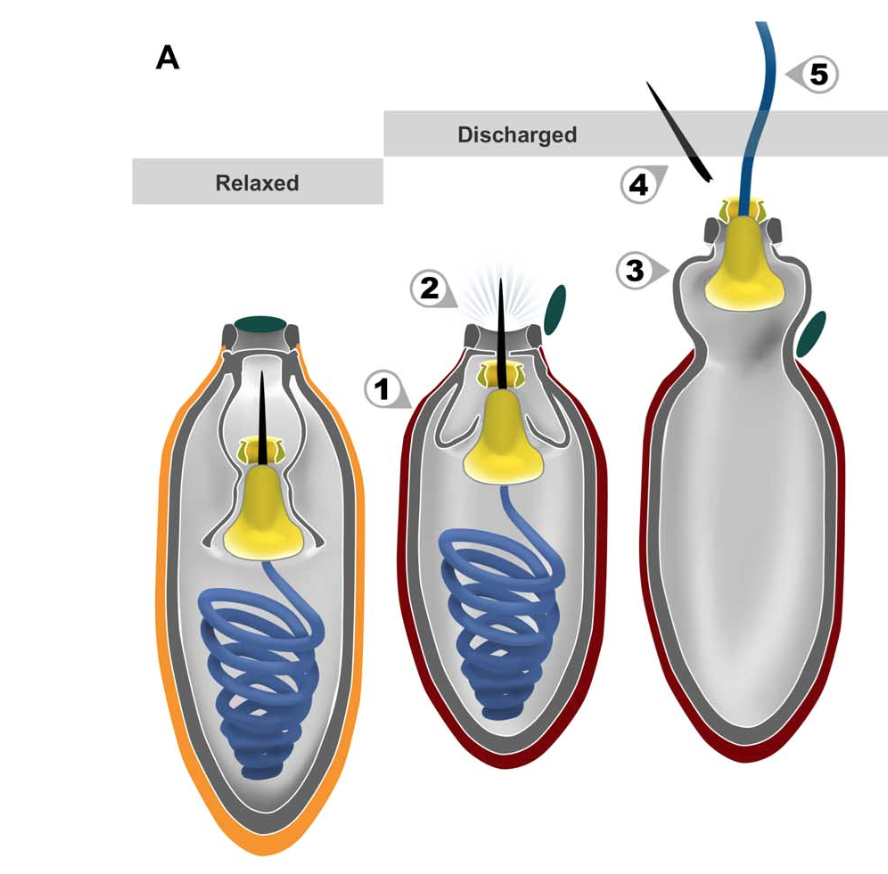
The putative ballistic sequence of nematocyst discharge is as follows:
(1) striated capsule contracts;
(2) internal pressure forces stylet to pierce capsule and open operculum (dark green);
(3) part of the capsule everts;
(4) stylet exits through the nozzle and detaches; and
(5) mucilaginous tubule (blue) uncoils and is projected through the stylet base (yellow). Orange, relaxed capsule wall; red, contracted capsule wall.
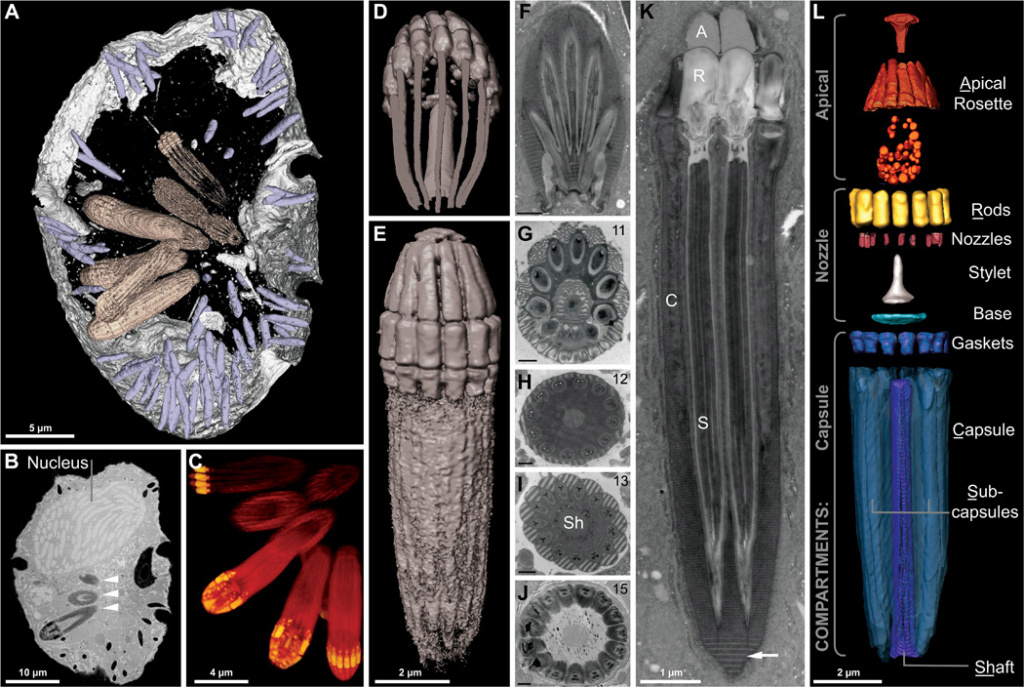
Reconstruction of the nematocysts in Nematodinium sp.
(A) Partial reconstruction of a cell with nematocysts shown in beige and mucocysts shown in purple.
(B) Longitudinal FIB-SEM section through a cell showing the nucleus and nematocysts (arrowheads).
(C) 3D maximum intensity projection showing a battery of nematocysts.
(D) 3D tilted reconstruction of a nematocyst.
(E) Longitudinal reconstruction of a nematocyst.
(F to K) Single-cell TEM micrographs of a nematocyst in oblique section
(F), cross section (G to J), and longitudinal section (K) showing the striated material (arrow) and variable symmetries across different nematocysts, with the number of subcapsules printed in the upper right corner. (L) A virtual dissection of the nematocyst components. Brackets indicate the membrane-bound compartments bounding the components indicated. (F) to (J) scale bar, 500 nm.
First Clear View of a One-Celled Harpooner in Action
https://www.youtube.com/watch?v=Otnw8qi8KaA&feature=youtu.be
First Clear View of a One-Celled Harpooner in Action7
Humans did not invent the harpoon gun. Some sea snails and jellyfish shoot darts at prey. And one single-celled predator even fires a projectile with a line attached so it can tow its prize away. Scientists working at the University of British Columbia reported Friday that they had captured the first high-resolution video of this microscopic harpooner in action. Although it is tempting to call it Ishmael, its real name is Polykrikos kofoidii. The video may lack the literary depth of “Moby Dick,” but it is strong on action, and Polykrikos is arguably better armed than any whaleboat launched from Captain Ahab’s Pequod. For example, it fires a projectile that has several working parts. First is an initial dart, called a taeniocyst. Gregory S. Gavelis, the primary author of a paper in Science Advances describing the research, said, “It just explodes.” Dr. Gavelis, now a researcher at Arizona State University, was pursuing his Ph.D. with Brian S. Leander when he did the work on Polykrikos. Dr. Leander and a group of other scientists contributed to the paper. Dr. Gavelis said the scientists did not yet know the function of the taeniocyst. “It might be a little glue bomb,” he said.
https://www.youtube.com/watch?v=CGq4KfyUtgk
After 5:00
In both cases this contractile capsule contracts after the upper Coulomb is dislodged and in addition in both cases this called up to Buell actually generates a lot of energy as it unfurls because it's actually somewhat rigid and it takes some energy to pack it into this organelle now this mechanistic force as a result of this coiled tubulin this contractile capsule combines with this sudden influx of water to accelerate this filament in some cases over 10,000 times the acceleration due to gravity now based on these similarities some scientists suggested that maybe these structures are related someway evolutionarily and so maybe these cnidarians acquired them from dinoflagellates or maybe both groups acquired them from bacteria or both groups developed them early on before they diverged way early on in evolutionary history in order to answer this more decisively scientists looked at this as polymer and more particularly the genes that code for the enzyme that produces this polymer in cnidarians and so they isolated these genes and they look for these genes or similar sequences in dinoflagellates and in other groups of life and what they found was that this polymer and the genes that produce this polymer are indeed found in bacteria and they seem to be homologous to the genes found in cnidarians areas now this suggests that these cnidarians did in fact acquire the ability to produce this osmotic propellant from bacteria but they didn't find was the same sets of genes or similar genes and dinoflagellates and so while they do believe dinoflagellates use some kind of osmotic propellant in the same way cnidarians do dinoflagellates don't produce this poly gamma gamma and they don't even have the genes required to produce it what this suggests is that this is really a case of convergent evolution where both lineages ended up producing somewhat similar structures but that these structures aren't actually related evolutionarily and this really isn't that surprising when you look at the fact that these new nematocysts are actually formed of multiple membrane-bound compartments and these organelle complexes and are actually quite a bit more complex structurally than these cnidarian nematocysts and so this is one case where a single celled protists was able to exceed the complexity at least in some regard that you see in multicellular animals
The researchers did a genetic analysis to try to understand how these structures evolved and see if they made their way into distant relatives, cnidarians, some of whom use similar ballistic weapons. This group includes sea anemones, jellyfish, box jellies, and Hydrozoa, including the Portuguese man o' war ( Physalia physalis ) . But, after sifting through the genetic blueprints of more than a hundred dinoflagellates, the researchers didn’t see any evidence that the weapons were shared or related to those used by cnidarians. Instead, the dinoflagellates seem to have developed their weapons independently.
Together, the authors conclude: “Despite the misconception that phytoplankton are passive cells, eukaryotic algae have given rise to (and arose from) multiple predatory lineages and, in the process, have independently evolved sophisticated ballistic organelles that exceed those of animals in complexity.
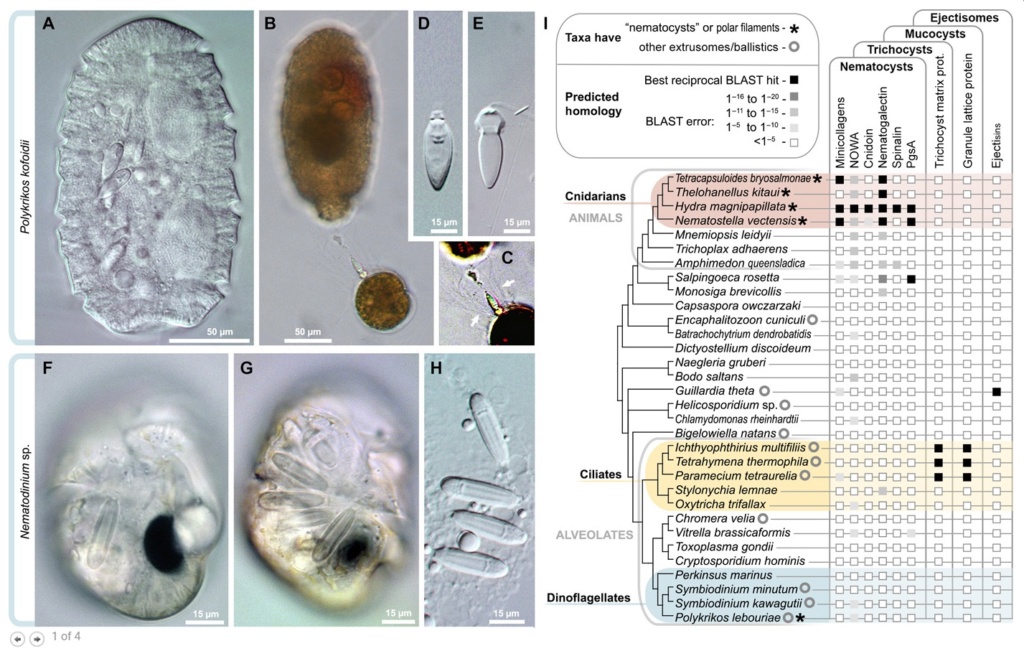
Diversity and independent origins of extrusomes.
(A to E) Nematocysts in the dinoflagellate P. kofoidii, including a live whole cell
(A) and a cell that was preserved in Lugol’s iodide solution while capturing a prey cell of A. tamarense
(B). (C) Enhanced contrast image shows the defensive trichocysts deployed by A. tamarense (arrows) in response to attack by P. kofoidii.
(D and E) Isolated nematocysts from P. kofoidii, seen as unfired (D) and discharged (E).
(F to H) Nematocysts in the dinoflagellate Nematodinium sp., which have an eye-like ocelloid (F). A battery of nematocysts is visible in the live cell (G) and remains intact after cell lysis (H).
(I) Genomic distribution of known extrusome proteins (vertical labels) across eukaryotes based on best reciprocal Basic Local Alignment Search Tool (BLAST) hits (black squares), which is a common predictor of protein homology. Taxa are listed within an established phylogenetic framework.
Convergence, another problem for evolution
Anemones, and jellyfish have morphologically (structurally) similar nematocysts as dinoflagellates, and yet are genetically distinct. An evolutionary interpretation of this data, then, demands that the morphologically identical organisms must have evolved independently of one another in a “repeatable” fashion. And this is a serious challenge and problem for evolutionary thinking.
Stephen J. Gould wrote in the book: Wonderful Life: The Burgess Shale and the Nature of History
“…No finale can be specified at the start, none would ever occur a second time in the same way, because any pathway proceeds through thousands of improbable stages. Alter any early event, ever so slightly, and without apparent importance at the time, and evolution cascades into a radically different channel.1
Gould’s metaphor of “replaying life’s tape” asserts that if one were to push the rewind button, erase life’s history, and let the tape run again, the results would be completely different. The very essence of the evolutionary process renders evolutionary outcomes nonreproducible and nonrepeatable. Therefore, “repeatable” evolution is inconsistent with the mechanism available to bring about biological change.
And Paleontologist J. William Schopf, one of the world’s leading authorities on early life on Earth, has made this very point in the book Life’s Origin.
Because biochemical systems comprise many intricately interlinked pieces, any particular full-blown system can only arise once…Since any complete biochemical system is far too elaborate to have evolved more than once in the history of life, it is safe to assume that microbes of the primal LCA cell line had the same traits that characterize all its present-day descendents.
https://www.youtube.com/watch?v=5BeqDD23oG4
After 7:00
https://www.youtube.com/watch?v=6VAp7DHut_E
https://www.youtube.com/watch?v=6zJiBc_N1Zk
1. https://facultyopinions.com/prime/727556446
2. https://soylentnews.org/article.pl?sid=17%2F04%2F06%2F1657216&from=rss&fbclid=IwAR0PUbd5rZY4vgqjxnI4P_PdqDei5wsCt9h_bHHNexSIhX7q2UO3pnAIF0E
3. https://arstechnica.com/science/2017/04/watch-militarized-microbes-use-some-sophisticated-weapons-to-snare-prey/
4. https://medium.com/science-and-philosophy/ocelloids-the-eyes-of-the-carnivorous-algae-18307c8a69f6
5. https://www.nature.com/articles/eye2017226
6. https://www.newscientist.com/term/evolution-of-the-eye/
7. https://www.nytimes.com/2017/03/31/science/dinoflagellate-video-one-celled-harpooner.html
8. https://www.livescience.com/58533-plankton-ballistic-weaponry-revealed.html
Two weird dinoflagellate protists have ballistic "nematocyst" organelles. The weaponry is complicated, savage, and unlike what’s seen in animals. These bugs are basically the battle tanks of the microbial world, with armor-piercing, multi-barrel Gatling guns for taking out prey. So cool! 1
Dinoflagellates called Nematodinium and Polykrikos are microscopic plankton, the kind of flotsam that whales gulp up by the ton. But these eukaryotic dinoflagellates, a type of protist, have their own drama-filled lives. They are hunters that eat other dinoflagellates, which themselves are bristling with armor, microscopic munitions and even chemical weapons. The tiny weapons of Nematodinium and related dinoflagellates are a unique invention: Though the weapons look a lot like the stingers of jellyfish, the structures are different. 8
Dinoflagellates are the coolest cells. Warnowiaceae dinoflagellates which also have nematocysts throwing out prey-capturing filaments are among the most complex single-celled organisms we know of. 4
Nematodinium's weird clusters of weaponry are called nematocysts. Gavelis and his colleagues used scanning electron microscopy (SEM) to image these nematocysts as well as the nematocysts of another well-armed dinoflagellate, Polykrikos kofoidii. They also captured the firing of P. kofoidii's weaponry in high-speed video for the first time.
What they saw were some intricately complex structures: Nematodinium has nematocysts that are clustered in rosette shapes and look like the multibarrel configuration of a Gatling gun.
Nobody has ever seen these creatures hunt, because Nematodinium is super-rare. "Unlike a Gatling gun, it probably shoots all of those capsules off at the same time."
Polykrikos kofoidii is a dinoflagellate that eats other dinoflagellates, capturing them with harpoon-like weapons that pierce the prey and drag them in to be consumed. This is a weird mechanism. This dinoflagellate sports capsules on its surface, each of which is topped by a finger-like projection called a taeniocyst. When it comes into contact with prey, the taeniocyst explodes, perhaps shooting out an adhesive similar to Spider-Man's webbing, Gavelis said. The eruption of the taeniocyst, in turn, triggers the capsule, or nematocyst, to shoot out a coiled tube tipped with a pointy, dagger-like projection called a stylet. The stylet pierces its way out of the capsule and penetrates the prey. The coil dissolves, but the stylet is still attached to the predator dinoflagellate by a towline. It uses that towline, basically like someone pulling a harpooned fish to its demise. Then, it's snack time.
Stinging sea animals, such as jellyfish and other cnidarians, have nematocysts, too. For that reason, there have long been theories that perhaps the dinoflagellates and the cnidarians evolved from some long-ago common ancestor with some pretty impressive defenses. Comparisons of the proteins found in cnidarians with those found in dinoflagellates indicates that there are no relationships.

A synthesis of fundamental differences between the nematocysts in cnidarians and dinoflagellates.
(a) A list of extrusomal features (with references given in 5, 22, 30, 31). (b) Illustrations of nematocyst development; tubules are orange, the dotted line in Nematodinium represents the plane of the cross section, on right. E = everting tubule. C = tiered compartments. S = subcapsules. (c) Illustrations of nematocysts firing. All illustrations are inferred from live observation except for that of Nematodinium, in which discharge has been only partially observed, in vitro. T = tow filament.
The genes that build nematocysts in dinoflagellates are entirely different from those that build them in cnidarians.
Biologists sometimes use the phrase "arms race" to describe a tug-of-war, but it's rarely this literal. Microbes called dinoflagellates have intricate weapons—including a microscopic version of a Gatling gun—to harpoon their dinners. Scientists have known about these harpoons for decades, and some have guessed that the weapons stem from the same source as the ones wielded by jellyfish and other cnidarians. An analysis of the genes and proteins involved with weapon construction, however, shows that dinoflagellates and cnidarians use different proteins to manufacture their weapons—meaning they arrived at similar solutions through separate paths. 2
Microbial arms race: Ballistic “nematocysts” in dinoflagellates represent a new extreme in organelle complexity
https://advances.sciencemag.org/content/3/3/e1602552
We examine the origin of harpoon-like secretory organelles (nematocysts) in dinoflagellate protists. These ballistic organelles have been hypothesized to be homologous to similarly complex structures in animals (cnidarians); but we show, using structural, functional, and phylogenomic data, that nematocysts
My comment: Why and how does that demonstrate a stepwise route ? It actually demonstrates the contrary. Namely that a stepwise route is not feasible because the structures are radically different.

Model of nematocyst homology and evolution in dinoflagellates.
(a) Illustration of homologous components in the known nematocysts of dinoflagellates as inferred from single-cell focused ion beam scanning electron microscopy (FIB-SEM) and serial TEM sections (15, 22, 30, 43, 44). Colors indicate homology of specific components of nematocysts (red = nozzle, blue = capsule, black = stylet, yellow = stylet base, purple = tubule, grey = uncertain), and horizontal grey lines indicate homology between the major membrane-bound compartments.
(b) Illustration of nematocyst traits mapped within a molecular phylogenetic context (based on fig. S6) showing the dinoflagellates with complex extrusomes. The number of major nematocyst vesicles/compartments is inferred to have increased over time along the lineage leading to polykrikoids (e.g., Polykrikos) and warnowiids (e.g., Nematodinium); the number of radially arranged subcompartments within nematocysts is inferred to have increased over time along the lineage leading to warnowiids (e.g., Nematodinium sp.).
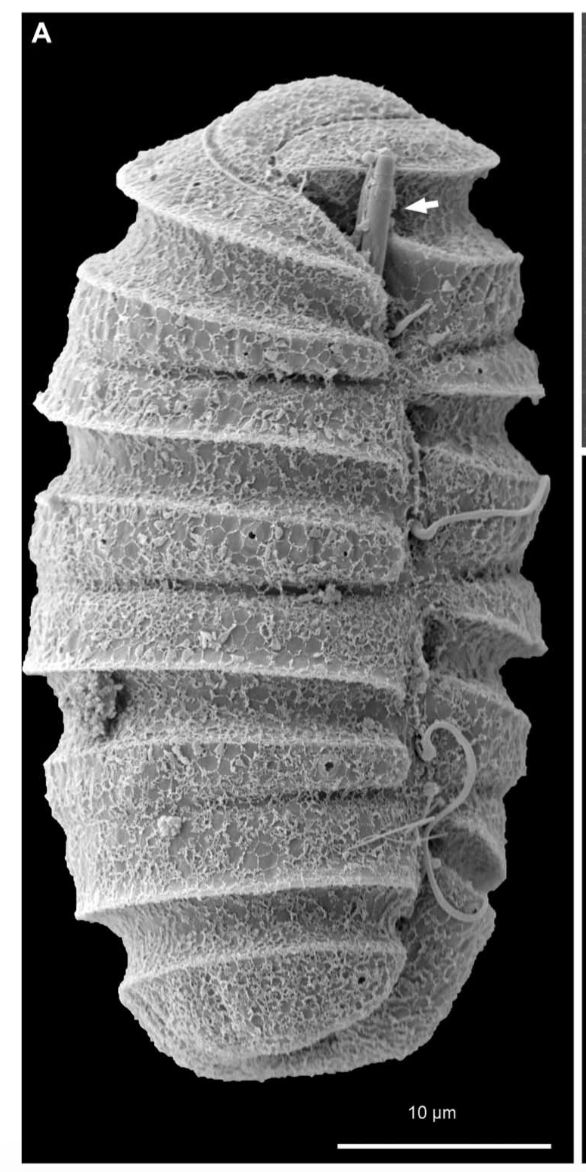
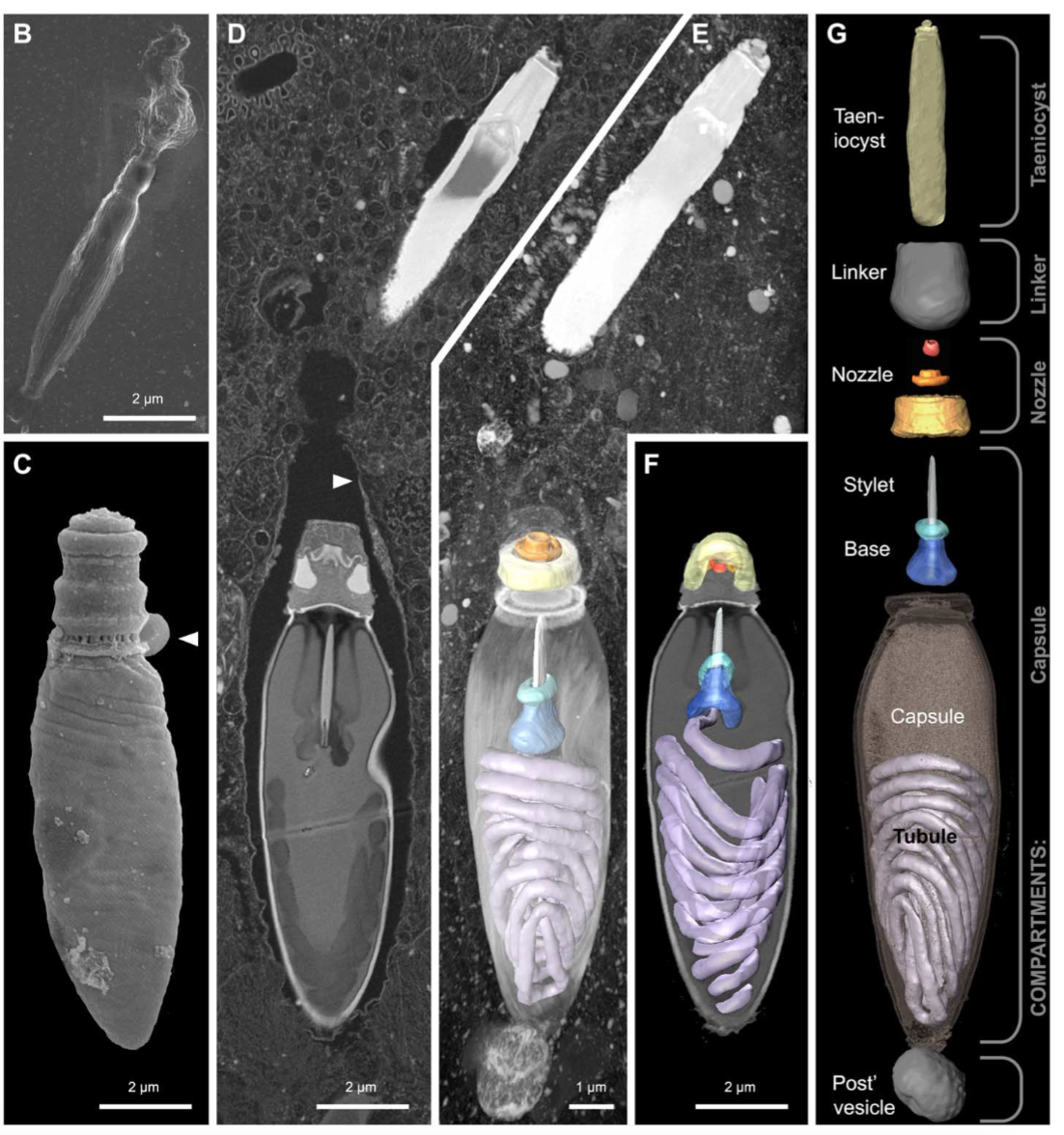
(B) SEM micrograph of an isolated taeniocyst that has discharged its amorphous contents.
(C) SEM micrograph of an isolated nematocyst that has become arrested very early in discharge; arrowhead, operculum.
(D) FIB-SEM section of taeniocyst (the white thing at the top) and nematocyst enclosed by a membranous chute (arrowhead) and
(E) maximum intensity projection of the same region seen slightly from above and below
(F). (G) Virtual dissection of the nematocyst-taeniocyst complex. Brackets indicate the membrane-bound compartments in which those components are grouped during early development. Later in development, compartments fuse to form the chute.
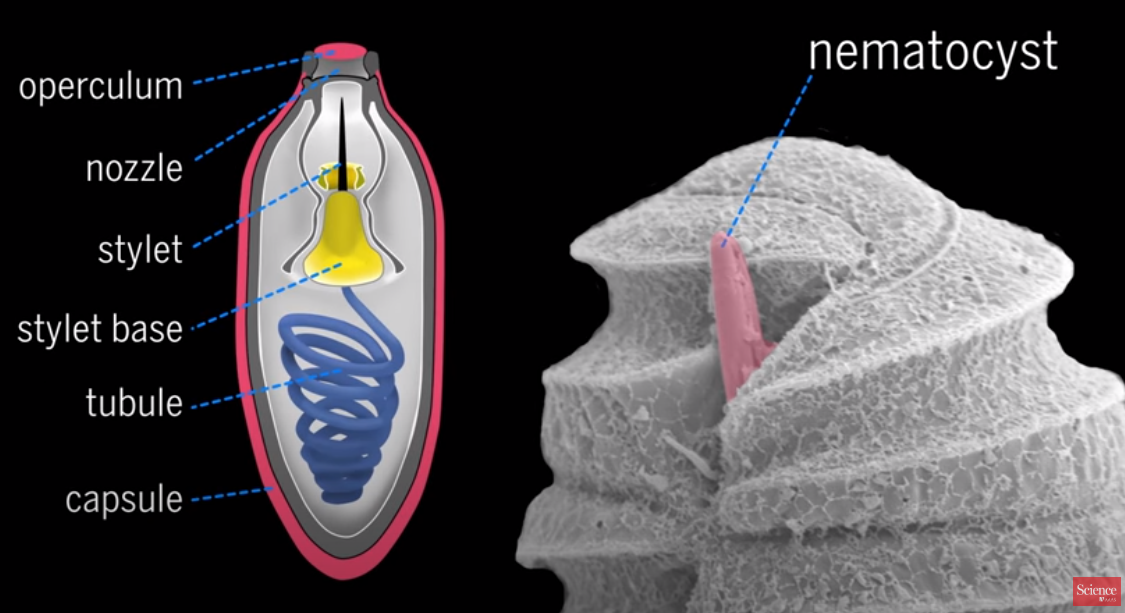
The capsule or nematocyst
1. Taeniocyst triggers the capsule, or nematocyst, to shoot out the coiled tube when it comes into contact with prey and explodes
2. a "linker" organelle. Use not known
3. a Nozzle designed to control the direction or characteristics of the tube flow as it exits the enclosed chamber
4. a base is where the tubule uncoils and is projected through
5. the stylet is a pointy dagger-like projection which pierces its way out of the capsule and penetrates the prey
6. the coiled Tube is tipped with a pointy, dagger-like projection called a stylet.
7. the capsule is the container of the tubes
8. a towline is used to pull the harpooned prey to its demise.
9. The tow filament is on the proximal end of the nematocyst and likely originates from the posterior vesicle.
This is an integrated interdependent system, where all parts must be in place in order to reach the higher end, that is to catch prey.
This dinoflagellate sports capsules on its surface, each of which is topped by a finger-like projection called a taeniocyst. When it comes into contact with prey, the taeniocyst explodes, perhaps shooting out an adhesive similar to Spider-Man's webbing. The eruption of the taeniocyst, in turn, triggers the capsule, or nematocyst, to shoot out a coiled tube tipped with a pointy, dagger-like projection called a stylet. The stylet pierces its way out of the capsule and penetrates the prey. The coil dissolves, but the stylet is still attached to the predator dinoflagellate by a towline. It uses that towline, basically like someone pulling a harpooned fish to its demise. Then, it's snack time.
The taeniocyst—that emerges from a finger-like projection near the top of each Polykrikos cell. Although the taeniocyst makes first contact with the prey, its functions are unclear. We provide evidence that taeniocysts are also ballistic structures. On five occasions, we observed that taeniocysts violently discharge when isolated from the cell by launching their contents through an apical channel (Fig. below).

The taeniocyst and nematocyst seem to work in tandem, with
(i) the taeniocyst intially adhering to the prey, followed by
(ii) discharge of the nematocyst, which punctures the prey, and last
(iii), the prey is retrieved using a tow filament.
Each mature nematocyst-taeniocyst complex resides within a single membrane-bound compartment (for example, the “chute”)
A novel “linker” organelle. This cylindrical structure connected the taeniocyst and the nematocyst further indicating that these two extrusomes types are deployed in succession, in a coordinated ballistic response. No comparable arrangement is found in cnidarians.
The putative ballistic sequence of nematocyst discharge is as follows:
(1) striated capsule contracts;
(2) internal pressure forces stylet to pierce capsule and open operculum (dark green);
(3) part of the capsule everts;
(4) stylet exits through the nozzle and detaches; and
(5) mucilaginous tubule (blue) uncoils and is projected through the stylet base (yellow). Orange, relaxed capsule wall; red, contracted capsule wall.

Reconstruction of the nematocysts in Nematodinium sp.
(A) Partial reconstruction of a cell with nematocysts shown in beige and mucocysts shown in purple.
(B) Longitudinal FIB-SEM section through a cell showing the nucleus and nematocysts (arrowheads).
(C) 3D maximum intensity projection showing a battery of nematocysts.
(D) 3D tilted reconstruction of a nematocyst.
(E) Longitudinal reconstruction of a nematocyst.
(F to K) Single-cell TEM micrographs of a nematocyst in oblique section
(F), cross section (G to J), and longitudinal section (K) showing the striated material (arrow) and variable symmetries across different nematocysts, with the number of subcapsules printed in the upper right corner. (L) A virtual dissection of the nematocyst components. Brackets indicate the membrane-bound compartments bounding the components indicated. (F) to (J) scale bar, 500 nm.
First Clear View of a One-Celled Harpooner in Action
https://www.youtube.com/watch?v=Otnw8qi8KaA&feature=youtu.be
First Clear View of a One-Celled Harpooner in Action7
Humans did not invent the harpoon gun. Some sea snails and jellyfish shoot darts at prey. And one single-celled predator even fires a projectile with a line attached so it can tow its prize away. Scientists working at the University of British Columbia reported Friday that they had captured the first high-resolution video of this microscopic harpooner in action. Although it is tempting to call it Ishmael, its real name is Polykrikos kofoidii. The video may lack the literary depth of “Moby Dick,” but it is strong on action, and Polykrikos is arguably better armed than any whaleboat launched from Captain Ahab’s Pequod. For example, it fires a projectile that has several working parts. First is an initial dart, called a taeniocyst. Gregory S. Gavelis, the primary author of a paper in Science Advances describing the research, said, “It just explodes.” Dr. Gavelis, now a researcher at Arizona State University, was pursuing his Ph.D. with Brian S. Leander when he did the work on Polykrikos. Dr. Leander and a group of other scientists contributed to the paper. Dr. Gavelis said the scientists did not yet know the function of the taeniocyst. “It might be a little glue bomb,” he said.
https://www.youtube.com/watch?v=CGq4KfyUtgk
After 5:00
In both cases this contractile capsule contracts after the upper Coulomb is dislodged and in addition in both cases this called up to Buell actually generates a lot of energy as it unfurls because it's actually somewhat rigid and it takes some energy to pack it into this organelle now this mechanistic force as a result of this coiled tubulin this contractile capsule combines with this sudden influx of water to accelerate this filament in some cases over 10,000 times the acceleration due to gravity now based on these similarities some scientists suggested that maybe these structures are related someway evolutionarily and so maybe these cnidarians acquired them from dinoflagellates or maybe both groups acquired them from bacteria or both groups developed them early on before they diverged way early on in evolutionary history in order to answer this more decisively scientists looked at this as polymer and more particularly the genes that code for the enzyme that produces this polymer in cnidarians and so they isolated these genes and they look for these genes or similar sequences in dinoflagellates and in other groups of life and what they found was that this polymer and the genes that produce this polymer are indeed found in bacteria and they seem to be homologous to the genes found in cnidarians areas now this suggests that these cnidarians did in fact acquire the ability to produce this osmotic propellant from bacteria but they didn't find was the same sets of genes or similar genes and dinoflagellates and so while they do believe dinoflagellates use some kind of osmotic propellant in the same way cnidarians do dinoflagellates don't produce this poly gamma gamma and they don't even have the genes required to produce it what this suggests is that this is really a case of convergent evolution where both lineages ended up producing somewhat similar structures but that these structures aren't actually related evolutionarily and this really isn't that surprising when you look at the fact that these new nematocysts are actually formed of multiple membrane-bound compartments and these organelle complexes and are actually quite a bit more complex structurally than these cnidarian nematocysts and so this is one case where a single celled protists was able to exceed the complexity at least in some regard that you see in multicellular animals
The researchers did a genetic analysis to try to understand how these structures evolved and see if they made their way into distant relatives, cnidarians, some of whom use similar ballistic weapons. This group includes sea anemones, jellyfish, box jellies, and Hydrozoa, including the Portuguese man o' war ( Physalia physalis ) . But, after sifting through the genetic blueprints of more than a hundred dinoflagellates, the researchers didn’t see any evidence that the weapons were shared or related to those used by cnidarians. Instead, the dinoflagellates seem to have developed their weapons independently.
Together, the authors conclude: “Despite the misconception that phytoplankton are passive cells, eukaryotic algae have given rise to (and arose from) multiple predatory lineages and, in the process, have independently evolved sophisticated ballistic organelles that exceed those of animals in complexity.

Diversity and independent origins of extrusomes.
(A to E) Nematocysts in the dinoflagellate P. kofoidii, including a live whole cell
(A) and a cell that was preserved in Lugol’s iodide solution while capturing a prey cell of A. tamarense
(B). (C) Enhanced contrast image shows the defensive trichocysts deployed by A. tamarense (arrows) in response to attack by P. kofoidii.
(D and E) Isolated nematocysts from P. kofoidii, seen as unfired (D) and discharged (E).
(F to H) Nematocysts in the dinoflagellate Nematodinium sp., which have an eye-like ocelloid (F). A battery of nematocysts is visible in the live cell (G) and remains intact after cell lysis (H).
(I) Genomic distribution of known extrusome proteins (vertical labels) across eukaryotes based on best reciprocal Basic Local Alignment Search Tool (BLAST) hits (black squares), which is a common predictor of protein homology. Taxa are listed within an established phylogenetic framework.
Convergence, another problem for evolution
Anemones, and jellyfish have morphologically (structurally) similar nematocysts as dinoflagellates, and yet are genetically distinct. An evolutionary interpretation of this data, then, demands that the morphologically identical organisms must have evolved independently of one another in a “repeatable” fashion. And this is a serious challenge and problem for evolutionary thinking.
Stephen J. Gould wrote in the book: Wonderful Life: The Burgess Shale and the Nature of History
“…No finale can be specified at the start, none would ever occur a second time in the same way, because any pathway proceeds through thousands of improbable stages. Alter any early event, ever so slightly, and without apparent importance at the time, and evolution cascades into a radically different channel.1
Gould’s metaphor of “replaying life’s tape” asserts that if one were to push the rewind button, erase life’s history, and let the tape run again, the results would be completely different. The very essence of the evolutionary process renders evolutionary outcomes nonreproducible and nonrepeatable. Therefore, “repeatable” evolution is inconsistent with the mechanism available to bring about biological change.
And Paleontologist J. William Schopf, one of the world’s leading authorities on early life on Earth, has made this very point in the book Life’s Origin.
Because biochemical systems comprise many intricately interlinked pieces, any particular full-blown system can only arise once…Since any complete biochemical system is far too elaborate to have evolved more than once in the history of life, it is safe to assume that microbes of the primal LCA cell line had the same traits that characterize all its present-day descendents.
https://www.youtube.com/watch?v=5BeqDD23oG4
After 7:00
https://www.youtube.com/watch?v=6VAp7DHut_E
https://www.youtube.com/watch?v=6zJiBc_N1Zk
1. https://facultyopinions.com/prime/727556446
2. https://soylentnews.org/article.pl?sid=17%2F04%2F06%2F1657216&from=rss&fbclid=IwAR0PUbd5rZY4vgqjxnI4P_PdqDei5wsCt9h_bHHNexSIhX7q2UO3pnAIF0E
3. https://arstechnica.com/science/2017/04/watch-militarized-microbes-use-some-sophisticated-weapons-to-snare-prey/
4. https://medium.com/science-and-philosophy/ocelloids-the-eyes-of-the-carnivorous-algae-18307c8a69f6
5. https://www.nature.com/articles/eye2017226
6. https://www.newscientist.com/term/evolution-of-the-eye/
7. https://www.nytimes.com/2017/03/31/science/dinoflagellate-video-one-celled-harpooner.html
8. https://www.livescience.com/58533-plankton-ballistic-weaponry-revealed.html
Last edited by Admin on Wed 21 Oct 2020 - 1:31; edited 6 times in total


