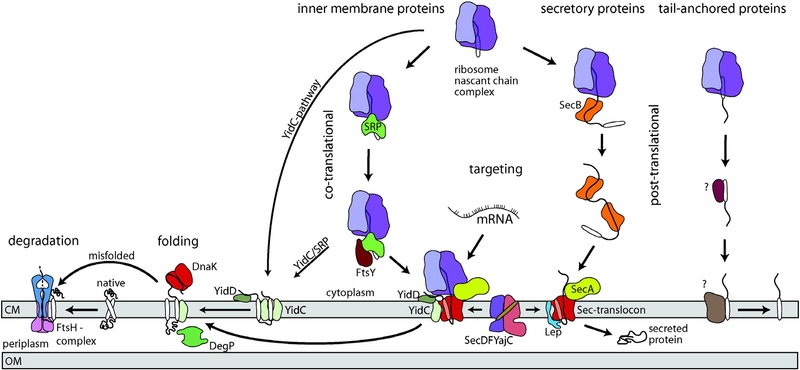
Assembly of bacterial inner membrane proteins 1
Many membrane proteins form multiple sub unit protein complexes. They possess integral and peripheral subunits. Enzymes known as Sec translocase and YidC insertase insert bacterial membrane proteins into the inner membrane. This process is assisted by YidC and the phospholipid phosphatidylethanolamine. Glycine zippers and other motifs also help transmembrane-transmembrane helix interactions that can form alpha helical bundles of membrane proteins. When membrane insertion occurs or when after membrane insertion occurs, the subunits of oligomeric membrane proteins have to be able to locate each other to construct the homo-oligiomeric and the hetero-oligomeric membrane complexes. Even though chaperones can serve as assembly factors to construct the oligomer, numerous protein oligomers seem to fold and oligomerize spontaneously. It has been shown by experiments that many of the subunits of hetero-olgiomers are structured after a sequential and patterned pathway to create the membrane protein complex. If it so happens that the inserted protein folds improperly or the membrane protein is assembled incorrectly, quality control mechanisms can deactivate the proteins.
Membrane protein can do a large variety of functions inside the cell from metabolite exchange to cell signaling and nerve conduction. They can also function as ATPases, electron carriers, ion channels, and transporters, sheddases, and photosynthetic reaction centers. They are abundant in both the eukaryotic and prokaryotic cell and they comprise about 20 percent to 30 percent of the total amount of proteins.
Many of the integral inner membrane proteins are alpha helical bundles with alpha helical membrane spanning areas. Advanced research has shown that the structures of the membrane proteins possess not only membrane spanning helices that are straight, but also possess very curved helices that span the membrane partially through. Alpha helical membrane proteins can exist as monomer or as multimeric complexes.
In order to guarantee that membrane proteins behave and function properly, they must be instructed to their destined membrane in the cell and then inserted and folded to the appropriate structure. Membrane tageting in the eukaryotic cells is necessary and more complicated than in eubacteria. Eukaryotic cells must instruct at least 10 membranes while eubacteria must only instruct 1 or 2 membranes in the gram-positive and gram-negative bacteria, respectively. After targeting, membrane protein integration and topogensis are instructed by a coordinated process of topogenic sequences and translocases. While this process is occurring, the transmembrane segments and extramembranous loops are folded.
The process of bacterial inner membrane protein assembling into the membrane is very complex. In addition, the mechanisms that control the protein targeting and inserstion into the membrane, folding of the alpha helical bundles, and the assembly into oligomeric membrane protein complexes will be explored more in depth.
The targeting of nascent chains to the membrane happens initially during the protein synthesis. It happens very early, even before the appearance of the polypeptide from the ribosomes channel. These nascent chains can already send signals in the ribosomes, which is a requirement of the signal recognition particle. A signal recognition particle is made up of a protein component Fth and a 4.5S RNA. The SRP combines with a hydrophobic part of a membrane protein as comes out from the ribosome at the membrane surface. The SRP-interacting area is most commonly the first TM region, but it can also be further apart and distinct from the TM segments. By studying the structure, it has been shown that a groove in the SRP M domain binds to the apolar segment.
Recognition and Targeting
When the receptor FTsy of the SRP- ribosome nascent chain complex is targeted by this complex, a SRP/FTsy complex is formed. The deconstruction of the complex and the freeing of the targeted protein needs GTP hydrolysis. The SRP and the FSty start out GTP bound and afterwards they construct into a complex by the interaction of their NG domains. A common trait between Ffh and FTsy is that they both have two homologous doamins and a distinct domain. By analyzing the structure if the Ffh and the FtsY NG doman complex, an interesting thing was found that there is a shared composite active-site area in the Ffh/FtsY hetereodimer, which is combined with two bound nucleotides. After the process of GTP hydrolysis, the membrane protein-nascent chain complex is sent to the SecYEG translocation channel, and the SRP and FtsY break apart from each other, which enables the SRP to recycl and interact in another round of SRP targeting. This sending of the nascent chain to the translocation channel is assisted by the interaction of the FtsY with SecY.
Insertion of the membrane proteins
It is necessary for the enzymes Translocases and intertases to put the freshly synthesized proteins into membranes. In bacteria, the SecYEG translocase and the YidC insertase have been depicted and analyzed. It reveals that they both display their translocation and insertion function in reconstituted systems. In addition, they are necessary processes for the bacterial life.
Sec Translocase Complex
The enzyme Sec translocase catalyzes the bacterial membrane protein insertion. The Sec translocase is made up of the membrane-embedded SeYEG and SecDFyajC complexes, in addition to the peripheral membrane component SecA. SecYEG supplies the protein-conducting channel. This is necessary for translocation and to make membrane protein insertion more efficient. Sec, which also known as the motor ATPase, is crucial for the translocation of preproteins through the membrane and for the translocation of particular hydrophilic areas of the membrane proteins. SecA utilizes ATP hydrolysis to propel the inserting polypeptide chain thorugh the Sec channel 20 to 30 residues simultaneously.
A major important discovery in the protein export area of studies was that the structure of the SecY complex was determine from an enzyme called Methanoccoccus jannaschii. This enzyme is made up of SecYEBeta. SecBeta does not have sequence homology to the eubacterial SecG but it does have sequence homology to the eukaryotic Sec61Beta. The SecY channel contains an hourglass structure with hydrophoibic narrow parts that is about 3 to 5 A in size which is found in the center of the channel. The narrow constriction wihtin the SecYEbeta splits the interior hydrophilic cavities on the periplasmic and cytoplasmic areas of the membrane. This narrow area is made up of a hydrophobic pore ring, which consists of 4 isoleucine residues, one valine, and one leucine residue. In addition, the aliphatic side chains of these amino acids are directed toward each other, which creates a hydrophobic collar through which the hydrophilic region of the polypeptide chain would be transport during translocation across the membrane.
Based on the crystal structure, the SecY channel is in sealed off state with the pore ring closed off by a helix on the luminal side. When the Sec channel opens up through signal peptide binding to the SecY TM2-TM7 region, the plug is relocated out of the channel site about 20 A away near the SecE helix.
Another important aspect of the SecY channel is the lateral gate. This is made to let the Tm regions of the inserting membrane proteins to be freed from the channel laterally and to split it into the lipid phase. The lateral gate is at the surface of SecY TM2 and TM7 of the Sec61alpha (SecY) which is found at the front side of the Sec channel. Before, TM2 and TM7 of the Sec61Alpha was thought to form the signal peptide-binding region because a signal peptide of preprotein can potentially be cross linked to these Tm parts during posttranslational translocation. When translocation of a polypeptide chain occurs, the lateral gate is opened up. The opening of this lateral gate is significant because locking the lateral gate by disulfide cross linking does not allow SecA-mediated preprotein translocation in Escherichia coli.
It is important to understand how the SecA operates with the SecY channel to translocate hydrophilic domains of membrane proteins across the membrane. The 4.5 A structure of the SecA/SecYEG from Thermotoga martima helps explain this process. First one copy of the SecA is attached to one copy of the SecY channel in the structure. The SecA is placed flat on the SecY channel about parallel to the membrane surface. It is important to note that the opening of the SecYEG channel has a two helix finger domain of SecA that can serve to transport substrates into the channel.
YidC Insertase
The YidC insertase is important because its job is to fit tiny proteins into the membrane. It was discovered that YidC influences membrane protein insertion. When the amounts of YidC is lessened in the cell, the insertion of Sec-independent proteins were slowed and discouraged. Before it was thought to be fit into the membrane spontaneously.
Through experiments it was thought that YidC affects the process of insertion of Sec-independent substrates. Photocross-linking studies that utilize a cell-free system displayed that membrane proteins that were stuck at different points of membrane protein insertion interact with YidC. Lipid vesicles that have YidC are enough to put the Sec-independent Pf3 coat protein and the ATP synthase subunit c. It was found that the Pf3 coat proteins sticks to the YiDC. This leads a significant conformational structure difference in the YidC protein.
1. https://en.wikibooks.org/wiki/Structural_Biochemistry/Proteins/Assembly_of_bacterial_inner_membrane_proteins
Many membrane proteins form multiple sub unit protein complexes. They possess integral and peripheral subunits. Enzymes known as Sec translocase and YidC insertase insert bacterial membrane proteins into the inner membrane. This process is assisted by YidC and the phospholipid phosphatidylethanolamine. Glycine zippers and other motifs also help transmembrane-transmembrane helix interactions that can form alpha helical bundles of membrane proteins. When membrane insertion occurs or when after membrane insertion occurs, the subunits of oligomeric membrane proteins have to be able to locate each other to construct the homo-oligiomeric and the hetero-oligomeric membrane complexes. Even though chaperones can serve as assembly factors to construct the oligomer, numerous protein oligomers seem to fold and oligomerize spontaneously. It has been shown by experiments that many of the subunits of hetero-olgiomers are structured after a sequential and patterned pathway to create the membrane protein complex. If it so happens that the inserted protein folds improperly or the membrane protein is assembled incorrectly, quality control mechanisms can deactivate the proteins.
Membrane protein can do a large variety of functions inside the cell from metabolite exchange to cell signaling and nerve conduction. They can also function as ATPases, electron carriers, ion channels, and transporters, sheddases, and photosynthetic reaction centers. They are abundant in both the eukaryotic and prokaryotic cell and they comprise about 20 percent to 30 percent of the total amount of proteins.
Many of the integral inner membrane proteins are alpha helical bundles with alpha helical membrane spanning areas. Advanced research has shown that the structures of the membrane proteins possess not only membrane spanning helices that are straight, but also possess very curved helices that span the membrane partially through. Alpha helical membrane proteins can exist as monomer or as multimeric complexes.
In order to guarantee that membrane proteins behave and function properly, they must be instructed to their destined membrane in the cell and then inserted and folded to the appropriate structure. Membrane tageting in the eukaryotic cells is necessary and more complicated than in eubacteria. Eukaryotic cells must instruct at least 10 membranes while eubacteria must only instruct 1 or 2 membranes in the gram-positive and gram-negative bacteria, respectively. After targeting, membrane protein integration and topogensis are instructed by a coordinated process of topogenic sequences and translocases. While this process is occurring, the transmembrane segments and extramembranous loops are folded.
The process of bacterial inner membrane protein assembling into the membrane is very complex. In addition, the mechanisms that control the protein targeting and inserstion into the membrane, folding of the alpha helical bundles, and the assembly into oligomeric membrane protein complexes will be explored more in depth.
The targeting of nascent chains to the membrane happens initially during the protein synthesis. It happens very early, even before the appearance of the polypeptide from the ribosomes channel. These nascent chains can already send signals in the ribosomes, which is a requirement of the signal recognition particle. A signal recognition particle is made up of a protein component Fth and a 4.5S RNA. The SRP combines with a hydrophobic part of a membrane protein as comes out from the ribosome at the membrane surface. The SRP-interacting area is most commonly the first TM region, but it can also be further apart and distinct from the TM segments. By studying the structure, it has been shown that a groove in the SRP M domain binds to the apolar segment.
Recognition and Targeting
When the receptor FTsy of the SRP- ribosome nascent chain complex is targeted by this complex, a SRP/FTsy complex is formed. The deconstruction of the complex and the freeing of the targeted protein needs GTP hydrolysis. The SRP and the FSty start out GTP bound and afterwards they construct into a complex by the interaction of their NG domains. A common trait between Ffh and FTsy is that they both have two homologous doamins and a distinct domain. By analyzing the structure if the Ffh and the FtsY NG doman complex, an interesting thing was found that there is a shared composite active-site area in the Ffh/FtsY hetereodimer, which is combined with two bound nucleotides. After the process of GTP hydrolysis, the membrane protein-nascent chain complex is sent to the SecYEG translocation channel, and the SRP and FtsY break apart from each other, which enables the SRP to recycl and interact in another round of SRP targeting. This sending of the nascent chain to the translocation channel is assisted by the interaction of the FtsY with SecY.
Insertion of the membrane proteins
It is necessary for the enzymes Translocases and intertases to put the freshly synthesized proteins into membranes. In bacteria, the SecYEG translocase and the YidC insertase have been depicted and analyzed. It reveals that they both display their translocation and insertion function in reconstituted systems. In addition, they are necessary processes for the bacterial life.
Sec Translocase Complex
The enzyme Sec translocase catalyzes the bacterial membrane protein insertion. The Sec translocase is made up of the membrane-embedded SeYEG and SecDFyajC complexes, in addition to the peripheral membrane component SecA. SecYEG supplies the protein-conducting channel. This is necessary for translocation and to make membrane protein insertion more efficient. Sec, which also known as the motor ATPase, is crucial for the translocation of preproteins through the membrane and for the translocation of particular hydrophilic areas of the membrane proteins. SecA utilizes ATP hydrolysis to propel the inserting polypeptide chain thorugh the Sec channel 20 to 30 residues simultaneously.
A major important discovery in the protein export area of studies was that the structure of the SecY complex was determine from an enzyme called Methanoccoccus jannaschii. This enzyme is made up of SecYEBeta. SecBeta does not have sequence homology to the eubacterial SecG but it does have sequence homology to the eukaryotic Sec61Beta. The SecY channel contains an hourglass structure with hydrophoibic narrow parts that is about 3 to 5 A in size which is found in the center of the channel. The narrow constriction wihtin the SecYEbeta splits the interior hydrophilic cavities on the periplasmic and cytoplasmic areas of the membrane. This narrow area is made up of a hydrophobic pore ring, which consists of 4 isoleucine residues, one valine, and one leucine residue. In addition, the aliphatic side chains of these amino acids are directed toward each other, which creates a hydrophobic collar through which the hydrophilic region of the polypeptide chain would be transport during translocation across the membrane.
Based on the crystal structure, the SecY channel is in sealed off state with the pore ring closed off by a helix on the luminal side. When the Sec channel opens up through signal peptide binding to the SecY TM2-TM7 region, the plug is relocated out of the channel site about 20 A away near the SecE helix.
Another important aspect of the SecY channel is the lateral gate. This is made to let the Tm regions of the inserting membrane proteins to be freed from the channel laterally and to split it into the lipid phase. The lateral gate is at the surface of SecY TM2 and TM7 of the Sec61alpha (SecY) which is found at the front side of the Sec channel. Before, TM2 and TM7 of the Sec61Alpha was thought to form the signal peptide-binding region because a signal peptide of preprotein can potentially be cross linked to these Tm parts during posttranslational translocation. When translocation of a polypeptide chain occurs, the lateral gate is opened up. The opening of this lateral gate is significant because locking the lateral gate by disulfide cross linking does not allow SecA-mediated preprotein translocation in Escherichia coli.
It is important to understand how the SecA operates with the SecY channel to translocate hydrophilic domains of membrane proteins across the membrane. The 4.5 A structure of the SecA/SecYEG from Thermotoga martima helps explain this process. First one copy of the SecA is attached to one copy of the SecY channel in the structure. The SecA is placed flat on the SecY channel about parallel to the membrane surface. It is important to note that the opening of the SecYEG channel has a two helix finger domain of SecA that can serve to transport substrates into the channel.
YidC Insertase
The YidC insertase is important because its job is to fit tiny proteins into the membrane. It was discovered that YidC influences membrane protein insertion. When the amounts of YidC is lessened in the cell, the insertion of Sec-independent proteins were slowed and discouraged. Before it was thought to be fit into the membrane spontaneously.
Through experiments it was thought that YidC affects the process of insertion of Sec-independent substrates. Photocross-linking studies that utilize a cell-free system displayed that membrane proteins that were stuck at different points of membrane protein insertion interact with YidC. Lipid vesicles that have YidC are enough to put the Sec-independent Pf3 coat protein and the ATP synthase subunit c. It was found that the Pf3 coat proteins sticks to the YiDC. This leads a significant conformational structure difference in the YidC protein.
1. https://en.wikibooks.org/wiki/Structural_Biochemistry/Proteins/Assembly_of_bacterial_inner_membrane_proteins
Last edited by Admin on Sat Mar 11, 2017 5:35 am; edited 1 time in total