M.Syvanen (2012): The flow of genes between different species represents a form of genetic variation. When I first started writing in 1982 about the implications of horizontal gene transfer (HGT) for macroevolutionary trends, the existence of the phenomenon was not yet accepted, given that evidence came only from isolated examples of bacterial plasmid transfer events and a case involving a retrovirus in mammals. There was great explanatory power in a theory that incorporated the notion of horizontally transferred genes as a source of genetic variation upon which natural selection acts. Reports of HGT in nature trickled in, but investigators debated the significance of HGT and the methodology for identifying it. In fact, only in the past 14 years has the number of examples of naturally occurring HGT become too large to ignore. With the recent availability of genome sequence data, the pace of discovery has picked up and interest in the phenomenon has increased. We now know that the ability of genes to function perfectly well across species boundaries has resulted in a significant horizontal flow of genes. Whether the genes are transferred by transposons, viruses, bacteria, or other vectors, or perhaps through direct contact or initial hybridization-like events, the horizontal flow of genes is a part of the story of life. Although the phenomenon of HGT is now widely accepted, current theoretical constructs remain quite resistant to many of its deeper implications. The first area I touch upon concerns the role of phylogenetic trees as a model for biological history. A second and related area concerns the continued speculation about and search for what is called the last universal common ancestor (LUCA) and the evolutionary significance of the biological unities. A third question involves the rethinking of higher taxonomic nomenclature. The chaotic phylogenetic relationships among plants remain unresolved, and a consideration of horizontalgene flow could help solve the puzzle. 24
Shelly Hamilich (2022): Our results suggest that horizontal gene transfer between hosts and their microbiota is a significant and active evolutionary mechanism that contributed new traits to plants and their commensal microbiota. 26
Comment: Rightly, Coppedge points out that: Information shared is not the same as information innovated, nor is borrowing a book as difficult as writing one. 27
Rama P. Bhatia (2022): the fitness effects of horizontally transferred genes are highly dependent on the environment. 28
Comment: The same problem to natural selection applies to HGT: since environmental factors influencing fitness effects would have to be taken into consideration in order to measure/calculate the influence of HGT in fitness, and since that is a variable that is stochastic, and cannot be measured, it becomes de facto impossible to detect up to what degree HGT influences fitness in populations in their natural environment.
Irreducible complexity C.Darwin (1860): “If it could be demonstrated that any complex organ existed, which could not possibly have been formed by numerous, successive, slight modifications, my theory would absolutely break down. But I can find no such case.” 30
Irreducible complexity, a term popularized by Michael Behe in his infamous book Darwin's Black box falsifies the claim that evolution explains the origin of biocomplexity and organismal form. If natural selection makes intelligent design superfluous, is it capable of instantiating proteins, cell types, organs, and organ systems, that have only function in cooperation/joint venture with other functional parts of the organism, or the organism as a whole, by slight, successive modifications over time? Does it select for structures, functions, traits, or what?
Behe, Darwin's Black Box (1996), page 39: By irreducibly complex I mean a single system composed of several well-matched, interacting parts that contribute to the basic function, wherein the removal of any one of the parts causes the system to effectively cease functioning. An irreducibly complex system cannot be produced directly (that is, by continuously improving the initial function, which continues to work by the same mechanism) by slight, successive modifications of a precursor system, because any precursor to an irreducibly complex system that is missing a part is by definition nonfunctional. An irreducibly complex biological system, if there is such a thing, would be a powerful challenge to Darwinian evolution.22
An irreducibly complex system is characterized by five points:1. a single system composed of several well-matched, interacting parts2. that contribute to the basic function3. the removal of any one of the parts causes the system to effectively cease functioning4. An irreducibly complex system cannot be produced directly (that is, by continuously improving the initial function, which continues to work by the same mechanism) by slight, successive modifications of a precursor system5. any precursor to an irreducibly complex system that is missing a part is by definition nonfunctional.1. Robert Carter: Genetic entropy and simple organisms 25 October 2012
2. Paul R. Ehrlich Natural Selection 19883.
Natural History Museum: What is natural selection?4. David Stack: Charles Darwin: Theory of Natural Selection 01 January 2021
5. Ernst Mayr: WHAT EVOLUTION IS A Conversation With Ernst Mayr [12.31.99]
6. J.Dekker: Natural Selection and its Four Conditions 2007
7. S.El-Showk: Natural selection: On fitness 2012
8. Evolution.Berkley: Evolutionary fitness
9. Adam Eyre-Walker: The distribution of fitness effects of new mutations August 2007
10. R. G. Brajesh: Distribution of fitness effects of mutations obtained from a simple genetic regulatory network model 08 July 2019
11. Thomas Bataillon: Effects of new mutations on fitness: insights from models and data 2014 Jul
18. Christopher J Graves: Variability in fitness effects can preclude selection of the fittest 2019 Sep 30
19. Vita Živa Alif: What is the best fitness measure in wild populations? A case study on the power of short-term fitness proxies to predict reproductive value November 19, 2021.
20. Ivana Cvijović: Fate of a mutation in a fluctuating environment August 24, 2015
21. Xia Hua: Darwinism for the Genomic Age: Connecting Mutation to Diversification 07 February 2017
22. Z Patwa: The fixation probability of beneficial mutations 29 July 2008
23. R. G. Brajesh: Distribution of fitness effects of mutations obtained from a simple genetic regulatory network model 08 July 2019
24. David F. Coppedge Evolutionary Fitness Is Not Measurable November 20, 2021
25. Michael Lynch: The frailty of adaptive hypotheses for the origins of organismal complexity May 15, 2007
26. Molly K Burke et.al.,: Genome-wide analysis of a long-term evolution experiment with Drosophila 2010 Sep 30
27. Ben Bradley: Natural selection according to Darwin: cause or effect? 11 April 2022
28. Adam Levy: How evolution builds genes from scratch 16 October 2019
29. J.Dulle: The (In)adequacy of Darwinian Evolution
30. Matthew Hurles: Gene Duplication: The Genomic Trade in Spare Parts July 13, 2004
31. Alisha K Holloway: Experimental evolution of gene duplicates in a bacterial plasmid model 2007 Feb
32. Joseph Esfandiar: Is gene duplication a viable explanation for the origination of biological information and complexity? 22 December 2010
33. Johan Hallin: Regulation plays a multifaceted role in the retention of gene duplicates November 22, 2019
34. Michael Lynch: The Origins of Genome Architecture 2007
35. Eugene V Koonin: Darwinian evolution in the light of genomics 2009 Mar
12. H. Allen Orr: Testing Natural Selection 2008
13. FRANCISCO J. AYALA: Darwin’s Greatest Discovery: Design Without Designer 2007
14.
Paul Gibson : Can Purifying Natural Selection Preserve Biological Information? – May 201315. Eugene V. Koonin :Toward a theory of evolution as multilevel learning February 4, 2022
16. Jerry A. Coyne, Why Evolution is True, p. 123. 2009
20. George Ellis: Controversy in Evolutionary Theory: A multilevel view of the issues. 2018
21. Armen Y. Mulkidjanian: Physico-Chemical and Evolutionary Constraints for the Formation and Selection of First Biopolymers: Towards the Consensus Paradigm of the Abiogenic Origin of Life 21 September 2007
22. M.Behe: Darwin's Black Box: The Biochemical Challenge to Evolution 1996
24. Michael Syvanen: Evolutionary Implications of Horizontal Gene Transfer 21 August 2012
25. Libretext: Horizontal Gene Transfer
26. Shelly Hamilich: Widespread horizontal gene transfer between plants and their microbiota August 26, 2022
27. David Coppedge: Gene Sharing Is More Widespread than Thought, with Implications for Darwinism September 20, 2022
28. Rama P. Bhatia: Environment and the Evolutionary Trajectory of Horizontal Gene Transfer April 01, 2022
30. Charles Darwin: Origin of Species: second British edition (1860), page 189
46. Ina Huang: [url=https://scholarblogs.emory.edu/evolutionshorts/2015/12/11/can-we-decide-the-direction-of-evolution/#:~:text=Many%20people%20believe%20that%20natural,random%20mutations%20that%20may%20occur
2Did life start polyphyletic, diversified, or monophyletic, with a universal common ancestor? Most, if not probably all science papers today start with the premise that universal common ancestry, and the tree of life, are true. Rather than questioning Darwin's hypothesis, 160 years old by now, it remains unquestioned. That is the basis upon which all evolutionary investigations today are made. It is accepted dogma and the starting point. The question asked is not: Is the tree of life true, and is universal common ancestry (UCA) true? But: How does the evidence fit into the tree? Even among some creationists, and intelligent design proponents, universal common ancestry is accepted as a plausible and justifiable possibility. But is it so? And what, when the nested hierarchy is not detectable, and evidence is lacking? it is just wiped under the table and ignored. There are many such cases. And scientific researchers find more and more such examples.
If one looks into the scientific literature, however, nothing is certain. O. Zhaxybayeva (2004): There is disagreement on the location of the root of the tree of life (e.g. different studies place the root:
1. within the bacterial domain or on the branch that leads to the bacterial domain;
2. within the eukaryotic domain;
3. within the archaeal domain;
4. yield inconclusive results. The timing of the organismal cenancestor is another unresolved question.41
In the meantime, the Genesis account of the special creation of each kind is often discarded from the onset or questioned. Some argue that Genesis does not require to be taken literally, it is perfectly fine to accept that it is an ancient near eastern myth, that uses figurative speech, and allegories that never were intended to be taken literally - so they say. Since Genesis is a several thousand-year-old book, a myth from ancient near eastern tribes, maybe even a copy from even older Mesopotamian texts, from the Babylonians, it by no means deserves to be trusted or taken literally. In special, when talking snakes, and angels with fiery, turning swords are mentioned. The Bible contains the highest information/semantic content in world literature. Genesis 1.1: "In the beginning, God created the heavens and the earth". That informs us in one short sentence about our origins. In information theory, semantics can be defined as the weight of the meanings” per sentence or per paragraph. There are thousands, maybe hundreds of thousands of books about origins, the beginning of the universe, life, and biodiversity, but none provide conclusive answers. All that scientific investigators can say in regard to our origins is: " probably, we suppose, we imagine, we theorize, we hypothesize, most likely, we suggest, it seems, it appears " etc. That extends through ALL evolutionary biology. The Bible, on the other hand, describes the origin of life in definitive terms: According to Genesis in the Bible, God created life during the creation week from the start and diversified it.
Gen. 1.11: Then God said, "Let the land produce vegetation: seed-bearing plants and trees on the land that bear fruit with seed in it, according to their various kinds." And it was so.
Gen. 1.21: So God created the great creatures of the sea and every living and moving thing with which the water teems, according to their kinds, and every winged bird according to its kind. And God saw that it was good.
Gen. 1.24: And God said, "Let the land produce living creatures according to their kinds: livestock, creatures that move along the ground, and wild animals, each according to its kind." And it was so.
Gen. 1.26/27: Then God said, "Let us make man in our image, in our likeness, and let them rule over the fish of the sea and the birds of the air, over the livestock, over all the earth, and over all the creatures that move along the ground." 27 So God created man in his own image, in the image of God he created him; male and female he created them.
It is important to outline: We have in Genesis a
complete account of origins. We can take two stances towards the Genesis account: 1. We believe it, or 2. We don't. Darwin contradicts the Genesis account, claiming that a Universal Common Ancestor existed, about 3,5 billion years ago, and gave rise to all biological forms and diversity. Which of the two accounts is most likely true?
Alberts (2022): The living world consists of three major divisions, or domains: eukaryotes, bacteria, and archaea. The great variety of living creatures that we see around us are eukaryotes. The name is from Greek, meaning “truly nucleated”, reflecting the fact that the cells of these organisms have their DNA enclosed in a membrane-bound organelle called the nucleus. Visible by simple light microscopy, this feature was used in the early twentieth century to classify living organisms as either eukaryotes (those with a nucleus) or prokaryotes (those without a nucleus). We now know that prokaryotes comprise two of the three major domains of life, bacteria, and archaea. Eukaryotic cells are typically much larger than those of bacteria and archaea; in addition to a nucleus, they typically contain a variety of membrane-bound organelles that are also lacking in prokaryotes. The genomes of eukaryotes also tend to run much larger—containing more than 20,000 genes for humans and corals, for example, compared with 4000–6000 genes for the typical bacteria or archaea. In addition to plants and animals, the eukaryotes include fungi (such as mushrooms or the yeasts used in beer- and bread-making), as well as an astonishing variety of single-celled, microscopic forms of life. 29
Was there a First and a Last Universal Common Ancestor?In his book, Darwin suggests that all living organisms are related by ascendency, and therefore they are all derived from ancestral species, which migrate around the world and diversify, generating the amazing biodiversity of organisms (Darwin, 1859).
K. Padian (2008): A sketch Darwin made soon after returning from his voyage on HMS Beagle (1831–36) showed his thinking about the diversification of species from a single stock (see Figure). This branching, extended by the concept of common descent, eventually formed an entire 'tree of life, developed enthusiastically by his German disciple Ernst Haeckel in the decades following the Origin. 1
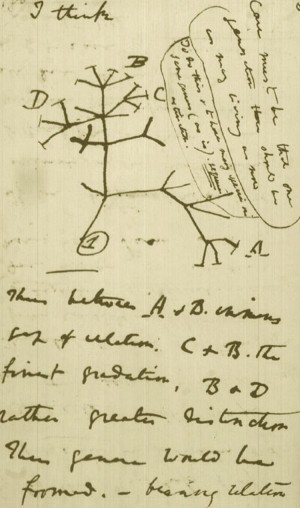
Charles Darwin's 1837 sketch of the diversification of species from a single stock. Credit: CAMBRIDGE UNIV. LIB./DARWIN-ONLINE.ORG.UK
D.Moran: Though the tree of life idea had been used to visualize taxonomy by Carl Linneaus, it became foundational as a tool for the development of Darwin’s evolutionary hypothesis. Lines connecting groups of organism branched off to more specific and supposedly related forms. Darwin saw that the connections made to groups and the position of species within a group were the result of shared similarities through ancestral descent. His theory was one attempt at explaining how those relationships might have come to exist. Ancestry was presumed to give rise to multiple lineages that diverged to create new life forms. Natural selection was the driving force for the divergence of species from a common ancestor. Natural variation within a type of organism was the generator of novel traits. Together, variation and selection would prove life evolved to its current time in existence.
M.A. Ragan (2009):
The rapid growth of genome-sequence data since the mid-1990s is now providing unprecedented detail on the genetic basis of life, and not surprisingly is catalyzing the most fundamental re-evaluation of origins and evolution since Darwin’s day. Several papers in this theme issue argue that Darwin’s tree of life is now best seen as an approximation—one quite adequate as a description of some parts of the living world (e.g. morphologically complex eukaryotes), but less helpful elsewhere (e.g. viruses and many prokaryotes); indeed, one of our authors goes farther, proclaiming the “demise” of Darwin’s tree as a hypothesis on the diversity and seeming naturalness of hierarchical arrangements of groups of living organisms. 16
10 reasons to refute the claim of Universal Common Ancestry 1. The DNA replication machinery is not homologous in the 3 domains of life. The bacterial core replisome enzymes do not share a common ancestor with the analogous components in eukaryotes and archaea. L.S. Kaguni (2016): Genome sequencing of cells from the three domains of life, bacteria, archaea, and eukaryotes, reveals that most of the core replisome components evolved twice, independently. Thus, the bacterial core replisome enzymes
do not share a common ancestor with the analogous components in eukaryotes and archaea, while the archaea and eukaryotic core replisome machinery share a common ancestor. An exception to this are the clamps and clamp loaders, which are homologous in all three domains of life.2
2. Bacteria and Archaea differ strikingly in the chemistry of their membrane lipids. There is no evidence of a common ancestor for any of the four glycolytic kinases or of the seven enzymes that bind nucleotides. S. Jain (2014): The composition of the phospholipid bilayer
is distinct in archaea when compared to bacteria and eukarya. In archaea, isoprenoid hydrocarbon side chains are linked via an ether bond to the sn-glycerol-1-phosphate backbone. In bacteria and eukarya on the other hand, fatty acid side chains are linked via an ester bond to the sn-glycerol-3-phosphate backbone. 3
Cell membrane phospholipids are synthesized by different, unrelated enzymes in bacteria and archaea, and yield chemically distinct membranes. Bacteria and archaea have membranes made of water-repellent fatty molecules. Bacterial membranes are made of fatty acids bound to the phosphate group while archaeal membranes are made of isoprenes bonded to phosphate in a different way. This leads to something of a paradox: Since a supposed last universal common ancestor, LUCA already had an impermeable membrane for exploiting proton gradients, why would its descendants have independently evolved two different kinds of impermeable membrane?Franklin M. Harold (2014):
Membranes also pose one of the most stubborn puzzles in all of cell evolution. Shortly after the discovery of the Archaea, it was realized that these organisms differ strikingly from the Bacteria in the chemistry of their membrane lipids. Archaea make their plasma membranes of isoprenoid subunits, linked by ether bonds to glycerol-1-phosphate; by contrast, Bacteria and Eukarya employ fatty acids linked by ester bonds to glycerol-3-phosphate. There are a few partial exceptions to the rule. Archaeal membranes often contain fatty acids, and some deeply branching Bacteria, such as Thermotoga, favor isoprenoid ether lipids (but even they couple the ethers to glycerol-3-phosphate). This pattern of lipid composition, which groups Bacteria and Eukarya together on one side and Archaea on the other, stands in glaring contrast to what would be expected from the universal tree, which puts Eukarya with the Archaea 143. Sequences of glycolytic enzymes differ between Archaea and Bacteria/Eukaryotes. S. F Alnomasy (2017): Some archaeal enzymes have some similarities with bacteria, but most archaeal enzymes have
no similarity with classical glycolytic pathways in Bacteria 4
There is no evidence of a common ancestor for any of the four glycolytic kinases or of the seven enzymes that bind nucleotides.Keith A. Webster (2003): There is no evidence of a common ancestor for any of the four glycolytic kinases or of the seven enzymes that bind nucleotides. Genetic, protein and DNA analysis, together with major differences in the biochemistry and molecular biology of all three domains – Bacteria, Archaea and Eukaryota – suggest that the three fundamental cell types are distinct and evolved separately (i.e. Bacteria are not actually pro-precursors of the eukaryotes, which have sequence similarities in particular parts of their biochemistry between both Bacteria or Archaea). Only a relatively small percentage of genes in Archaea have sequence similarity to genes in Bacteria or Eukaryota. Furthermore, most of the cellular events triggered by intracellular Ca2+ in eukaryotes do not occur in either Bacteria or Archaea. 154. There are at least six distinct autotrophic carbon fixation pathways.If common ancestry were true, an ancestral Wood–Ljungdahl pathway should have become life's one and only principle for biomass production. W. Nitschke (2013):
At least six distinct autotrophic carbon fixation pathways have been elucidated during the past few decades 5 Since the claim is that this is how life began fixing carbon, and the first carbon fixation pathways were anaerobic, this represents a major puzzle for proponents of common ancestry, and its proponents are led to wonder why an ancestral Wood–Ljungdahl pathway has not become life's one and only principle for biomass production. What is even more puzzling, is the fact that searches of the genomes of acetogenins for enzymes clearly homologous to those of the methanogenic C1-branch came up empty-handed with one notable exception, i.e. the initial step of CO2 reduction which is, in both cases, catalyzed by a molybdo/tungstopterin enzyme from the complex iron-sulfur molybdoenzyme (CISM) superfamily. So, partially, carbon fixation pathways share partially the same enzymes. This points clearly to a common designer choosing different routes for the same reaction but using partially convergent design. Similarities between living organisms could be because they have been designed by the same intelligence, just as we can recognize a Norman Foster building by his characteristic style, or a painting by Van Gogh. We expect to see repeated motifs and re-used techniques in different works by the same artist/designer.
5. There is a sharp divide in the organizational complexity of the cell between eukaryotes and prokaryotesE. V. Koonin (2010): There is a sharp divide in the organizational complexity of the cell between eukaryotes, which have complex intracellular compartmentalization, and even the most sophisticated prokaryotes (archaea and bacteria), which do not. The compartmentalization of eukaryotic cells is supported by an elaborate endomembrane system and by the actin-tubulin-based cytoskeleton. There are no direct counterparts of these organelles in archaea or bacteria. The other hallmark of the eukaryotic cell is the presence of mitochondria, which have a central role in energy transformation and perform many additional roles in eukaryotic cells, such as in signaling and cell death. 6
6. A typical eukaryotic cell is about 1,000-fold bigger by volume than a typical bacterium or archaeonE. V. Koonin (2010):
The origin of eukaryotes is a huge enigma and a major challenge for evolutionary biology. There is a sharp divide in the organizational complexity of the cell between eukaryotes, which have complex intracellular compartmentalization, and even the most sophisticated prokaryotes (archaea and bacteria), which do not. A typical eukaryotic cell is about 1,000-fold bigger by volume than a typical bacterium or archaeon, and functions under different physical principles: free diffusion has little role in eukaryotic cells, but is crucial in prokaryotes. The compartmentalization of eukaryotic cells is supported by an elaborate endomembrane system and by the actin-tubulin-based cytoskeleton. There are no direct counterparts of these organelles in archaea or bacteria. The other hallmark of the eukaryotic cell is the presence of mitochondria, which have a central role in energy transformation and perform many additional roles in eukaryotic cells, such as in signaling and cell death. 67. Horizontal gene transfer (HGT)E. V. Koonin (2012):
Subsequent massive sequencing of numerous, complete microbial genomes have revealed novel evolutionary phenomena, the most fundamental of these being: pervasive horizontal gene transfer (HGT), in large part mediated by viruses and plasmids, that shapes the genomes of archaea and bacteria and call for a radical revision (if not abandonment) of the Tree of Life concept 78. RNA Polymerase differencesRNA Polymerase differences: Prokaryotes only contain three different promoter elements: -10, -35 promoters, and upstream elements. Eukaryotes contain many different promoter elements: TATA box, initiator elements, downstream core promoter element, CAAT box, and the GC box to name a few. Eukaryotes have three types of RNA polymerases, I, II, and III, and prokaryotes only have one type. Eukaryotes form and initiation complex with the various transcription factors that dissociate after initiation is completed. There is no such structure seen in prokaryotes. Another main difference between the two is that transcription and translation occurs simultaneously in prokaryotes and in eukaryotes the RNA is first transcribed in the nucleus and then translated in the cytoplasm. RNAs from eukaryotes undergo post-transcriptional modifications including: capping, polyadenylation, and splicing. These events do not occur in prokaryotes. mRNAs in prokaryotes tend to contain many different genes on a single mRNA meaning they are polycystronic. Eukaryotes contain mRNAs that are monocystronic. Termination in prokaryotes is done by either rho-dependent or rho-independent mechanisms. In eukaryotes transcription is terminated by two elements: a poly(A) signal and a downstream terminator sequence. 8
9. Ribosome and ribosome biogenesis differencesRibosome and ribosome biogenesis differences: Although we could identify E. coli counterparts with comparable biochemical activity for 12 yeast ribosome biogenesis factors (RBFs), only 2 are known to participate in bacterial ribosome assembly. This indicates that the recruitment of individual proteins to this pathway has been largely independent in the bacterial and eukaryotic lineages. The bacterial version of a universal ribosomal protein tends to be remarkably different from its archaeal equivalent, the same being true, even more dramatically, for the aminoacyl-tRNA synthetases. In both cases, in a sequence alignment, a position constant in composition in the Bacteria tends to be so in its archaeal homolog as well, but the archaeal and bacterial compositions for that position often differ from each other. Moreover, among the aminoacyl-tRNA synthetases, a total lack of homology between large (and characteristic) sections of the bacterial version of a molecule and its archaeal counterpart is common. 9 10. The replication promoters of bacterial and yeast genes have different structuresA. C. Leonard (2013):
Like the origins of DNA replication, the promoters of bacterial and yeast genes have different structures, are recognized by different proteins, and are not exchangeable. The absolute incompatibility between prokaryote (e.g., E. coli) and eukaryote (e.g., yeast) origins of replication and promoters, as well as DNA replication, transcription, and translation machineries, stands as a largely unrecognized challenge to the evolutionary view that the two share a common ancestor. 10
No common ancestor for VirusesEugene V. Koonin (2020): In the genetic space of viruses and MGEs, no genes are universal or even conserved in the majority of viruses. Viruses have several distinct points of origin, so there has never been a last common ancestor of all viruses. 11
Viruses and the tree of life (2009): Viruses are polyphyletic: In a phylogenetic tree, the characteristics of members of taxa are inherited from previous ancestors. Viruses cannot be included in the tree of life because they do not share characteristics with cells, and no single gene is shared by all viruses or viral lineages. While cellular life has a single, common origin, viruses are polyphyletic – they have many evolutionary origins. Viruses don’t have a structure derived from a common ancestor. Cells obtain membranes from other cells during cell division. According to this concept of ‘membrane heredity’, today’s cells have inherited membranes from the first cells. Viruses have no such inherited structure. They play an important role by regulating population and biodiversity. 12
Eugene V. Koonin (2017): The entire history of life is the story of virus–host coevolution. Therefore the origins and evolution of viruses are an essential component of this process. A signature feature of the virus state is the capsid, the proteinaceous shell that encases the viral genome. Although homologous capsid proteins are encoded by highly diverse viruses, there are at least 20 unrelated varieties of these proteins. Viruses are the most abundant biological entities on earth and show remarkable diversity of genome sequences, replication and expression strategies, and virion structures. Evolutionary genomics of viruses revealed many unexpected connections but the general scenario(s) for the evolution of the virosphere remains a matter of intense debate among proponents of the cellular regression, escaped genes, and primordial virus world hypotheses. A comprehensive sequence and structure analysis of major virion proteins indicates that they evolved on about 20 independent occasions. Virus genomes typically consist of distinct structural and replication modules that recombine frequently and can have different evolutionary trajectories. The present analysis suggests that, although the replication modules of at least some classes of viruses might descend from primordial selfish genetic elements, bona fide viruses evolved on multiple, independent occasions throughout the course of evolution by the recruitment of diverse host proteins that became major virion components. 13
Comment: The importance of the admission that viruses do not share a common ancestor cannot be outlined enough. Researchers also admit, that under a naturalistic framework, the origin of viruses remains obscure, and has not found an explanation. One reason is that viruses depend on a cell host in order to replicate. Another is, that the virus capsid shells that protect the viral genome are unique, there is no counterpart in life. A science paper that I quote below describes capsids with a "geometrically sophisticated architecture not seen in other biological assemblies". This seems to be interesting evidence of design. The claim that their origin has something to do with evolution is also misleading - evolution plays no role in explaining either the origin of life or the origin of viruses. The fact that "no single gene is shared by all viruses or viral lineages" prohibits drawing a tree of viruses leading to a common ancestor.
D M Raup (1983):
Life forms are made possible by the remarkable properties of polypeptides. It has been argued that there must be many potential but unrealized polypeptides that could be used in living systems. The number of possible primary polypeptide structures with lengths comparable to those found in living systems is almost infinite. This suggests that the particular subset of polypeptides of which organisms are now composed is only one of a great many that could be associated in viable biochemistries. There is no taxonomic category available to contain all life forms descended from a single event of life origin. Here, we term such a group, earthly or otherwise, a bioclade. If more than one bioclade survives, life is polyphyletic. If only one survives, it is monophyletic. We conclude that multiple origins of life in the early Precambrian is a reasonable possibility.21W. Ford Doolittle (2007):
Darwin claimed that a unique inclusively hierarchical pattern of relationships between all organisms based on their similarities and differences [the Tree of Life (TOL)] was a fact of nature, for which evolution, and in particular a branching process of descent with modification, was the explanation. However, there is no independent evidence that the natural order is an inclusive hierarchy, and incorporation of prokaryotes into the TOL is especially problematic. The only data sets from which we might construct a universal hierarchy including prokaryotes, the sequences of genes, often disagree and can seldom be proven to agree. Hierarchical structure can always be imposed on or extracted from such data sets by algorithms designed to do so, but at its base the universal TOL rests on an unproven assumption about pattern that, given what we know about process, is unlikely to be broadly true. This is not to say that similarities and differences between organisms are not to be accounted for by evolutionary mechanisms, but descent with modification is only one of these mechanisms, and a single tree-like pattern is not the necessary (or expected) result of their collective operation. Pattern pluralism (the recognition that different evolutionary models and representations of relationships will be appropriate, and true, for different taxa or at different scales or fordifferent purposes) is an attractive alternative to the quixotic pursuit of a single true TOL.20
Douglas L. Theobald (2010):
In all cases tried, with a wide variety of evolutionary models (from the simplest to the most parameter rich), the multiple-ancestry models for shuffled data sets are preferred by a large margin over common ancestry models (LLR on the order of a thousand), even with the large internal branches. 17C. P. Kempes (2021):
We argue for multiple forms of life realized through multiple different historical pathways. From this perspective, there have been multiple origins of life on Earth—life is not a universal homology. By broadening the class of originations, we significantly expand the data set for searching for life. We define life as the union of two crucial energetic and informatic processes producing an autonomous system that can metabolically extract and encode information from the environment of adaptive/survival value and propagate it forward through time. We provide a new perspective on the origin of life by arguing that life has emerged many times on Earth and that there are many forms of extant life coexisting on a variety of physical substrates. The ultimate theory of life will certainly have ingredients from abstract theories of engineering, computation, physics, and evolution, but we expect will also require new perspectives and tools, just as theories of computation have. It should be able to highlight life as the ultimate homoplasy (convergence) rather than homology, where life is discovered repeatedly from many different trajectories.A science forum was held at Arizona State University in February 2011, where the following dialogue between Dawkins and Venter was reported:Venter: I'm not so sanguine as some of my colleagues here that there's only one life form on this planet we have a lot of different types of metabolism different organisms I wouldn't call you the same life-form as the one we have that lives in pH12 base that would dissolve your skin if we drop you at it. The same genetic it will have a common anything well you don't have the same genetic code in fact the micoplasmas use a different genetic code and it would not work in yourself so there are a lot of variations on the unitDawkins: But you're not saying it belongs to a different tree of life from meVenter: I well I think the Tree of Life is an artifact of some early scientific studies that aren't really holding up so the tree you know there may be a bush of life. Bush I don't like that word written but that's only I can see that so there's not a tree of life and in fact from our deep sequencing of organisms in the ocean out of now we have about 60 million different unique gene sets we found 12 that looked like a very very deep branching perhaps fourth domain of life that obviously is extremely rare that it only shows up out of those few sequences but it's still DNA based but you know the diversity we have in the DNA world I'm not so saying what in wedding ready to throw out the DNA world. 18 19From the Last Universal Common Ancestor, LUCA, to Eukaryotic cellsC. Woese (2002): The evolution of modern cells is
arguably the most challenging and important problem the field of Biology has ever faced. 40
G. E. Mikhailovsky (2021): It is puzzling why life on Earth consisted of prokaryotes for up to 2.5 ± 0.5 billion years (Gy) before the appearance of the first eukaryotes. This period, from LUCA (Last Universal Common Ancestor) to LECA (Last Eucaryotic Common Ancestor), we have named the Lucacene, to suggest all prokaryotic descendants of LUCA before the appearance of LECA. The structural diversity of eukaryotic organisms is very large, while the morphological diversity of prokaryotic cells is immeasurably lower. 37
EukaryotesAs everywhere through evolutionary biology, the claim is that things went from less, to more complex, over long periods of time.
D. A. Peattie: The eukaryote has structural features that allow it to communicate better than prokaryotes, features that permit cellular aggregation and multicellular life. In contrast, the more primitive prokaryotes are less well-equipped for intercellular communication and cannot readily organize into multicellular organisms.
Not only do eukaryotic cells allow larger and more complex organisms to be made, but they are themselves larger and more complex than prokaryotic cells. Whether eukaryotic cells live singly or as part of a multicellular organism, their activities can be much more complex and diversified than those of their prokaryotic counterparts. In prokaryotes, all internal cellular events take place within a single compartment, the cytoplasm. Eukaryotes contain many subcellular compartments, called organelles. Even single-celled eukaryotes can display remarkable complexity of function; some have features as specialized and diverse as sensory bristles, mouth parts, muscle-like contractile bundles, or stinging darts.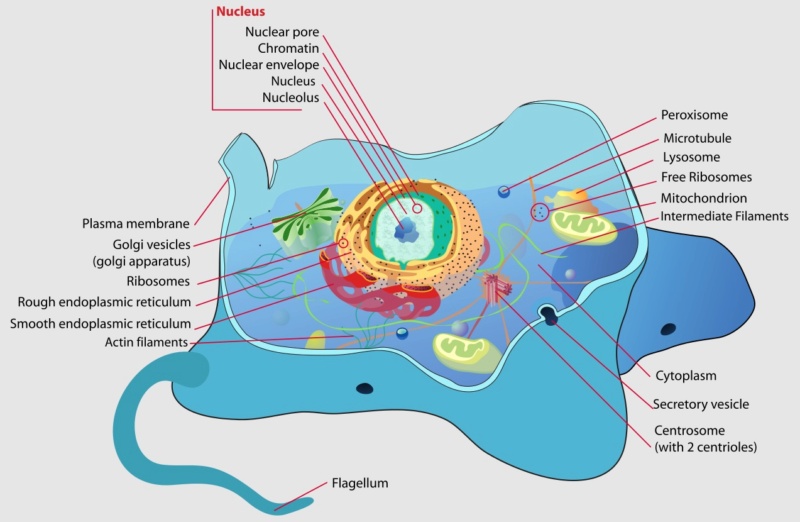
Structure of a typical animal cell
On a very fundamental level, eukaryotes and prokaryotes are similar. They share many aspects of their basic chemistry, physiology and metabolism. Both cell types are constructed of and use similar kinds of molecules and macromolecules to accomplish their cellular work. In both, for example, membranes are constructed mainly of fatty substances called lipids, and molecules that perform the cell's biological and mechanical work are called proteins.Eukaryotes and prokaryotes both use the same chemical relay system to make protein. A permanent record of the code for all of the proteins the cell will require is stored in the form of DNA. Because DNA is the master copy of the cell's (or organism's) genetic make-up, the information it contains is absolutely crucial to the maintenance and perpetuation of the cell. As if to safeguard this archive, the cell does not use the DNA directly in protein synthesis but instead copies the information onto a temporary template of RNA, a chemical relative of DNA. Both the DNA and the RNA constitute a "recipe" for the cell's proteins. The recipe specifies the order in which amino acids, the chemical subunits of proteins, should be strung together to make the functional protein. Protein synthesis both in eukaryotes and prokaryotes takes place on structures called ribosomes, which are composed of RNA and protein. This illustrates one way in which prokaryotes and eukaryotes are similar and highlights the idea that differences between these organisms are often architectural. In other words, both cell types use the same bricks and mortar, but the structures they build with these materials vary dramatically.The prokaryotic cell can be compared to a studio apartment: a one-room living space that has a kitchen area abutting the living room, which converts into a bedroom at night. All necessary items fit into their own locations in one room. There is an everyday; washable rug. Room temperature is comfortable-not too hot, not too cold. Conditions are adequate for everything that must occur in the apartment, but not optimal for any specific activity. In a similar way, all of the prokaryote's functions fit into a single compartment. The DNA is attached to the cell's membrane. Ribosomes float freely in the single compartment. Cellular respiration-the process by which nutrients are metabolized to release energy-is carried out at the cell membrane; there is no dedicated compartment for respiration. A eukaryotic cell can be compared to a mansion, where specific rooms are designed for particular activities. The mansion is more diverse in the activities it supports than the studio apartment. It can accommodate overnight guests comfortably and support social activities for adults in the living room or dining room, for children in the playroom. The baby's room is warm and furnished with bright colors and a soft, thick carpet. The kitchen has a stove, a refrigerator and a tile floor. Items are kept in the room that is most appropriate for them, under conditions ideal for the activities in that specific room.
A eukaryotic cell resembles a mansion in that it is subdivided into many compartments. Each compartment is furnished with items and conditions suitable for a specific function, yet the compartments work together to allow the cell to maintain itself, to replicate and to perform more specialized activities.Taking a closer look, we find three main structural aspects that differentiate prokaryotes from eukaryotes. The definitive difference is the presence of a true (eu) nucleus (karyon) in the eukaryotic cell. The nucleus, a double-membrane casing, sequesters the DNA in its own compartment and keeps it separate from the rest of the cell. In contrast, no such housing is provided for the DNA of a prokaryote. Instead the genetic material is tethered to the cell membrane and is otherwise allowed to float freely in the cell's interior. It is interesting to note that the DNA of eukaryotes is attached to the nuclear membrane, in a manner reminiscent of the attachment of prokaryotic DNA to the cell's outer membrane. 28The greatest discontinuity in evolution: The gap from prokaryotes to eukaryotes
Ro Y. STANIER et. al., (1963) “The basic divergence in cellular structure, which separates the bacteria and blue-green algae from all other cellular organisms, represents
the greatest single evolutionary discontinuity to be found in the presentday world” 31
E. V. Koonin (2002):
The eukaryotic chromatin remodeling machinery, the cell cycle regulation systems, the nuclear envelope, the cytoskeleton, and the programmed cell death (PCD, or apoptosis) apparatus all are such major eukaryotic innovations, which do not appear to have direct prokaryotic predecessors.25
E. Derelle et.al.,(2006):
The unicellular green marine alga Ostreococcus tauri is the world's smallest free-living eukaryote known to date, and encodes the fewest number of genes. It has been hypothesized, based on its small cellular and genome sizes, that it may reveal the “bare limits” of life as a free-living photosynthetic eukaryote, presumably having disposed of redundancies and presenting a simple organization and very little noncoding sequence. 27 It has a genome size of 12.560,000 base pairs, 8,166 genes and 7745 proteins. in comparison, the simplest free-living bacteria today is
Pelagibacter ubique get by with about 1,300 genes and 1,308,759 base pairs and code for 1,354 proteins.
T. Cavalier-Smith (2010): This radical transformation of cell structure (eukaryogenesis) is the most complex and extensive case of quantum evolution in the history of life. Beforehand earth was a sexless, purely bacterial and viral world. Afterwards sexy, endoskeletal eukaryotes evolved morphological complexity: diatoms, butterflies, corals, whales, kelps, and trees 32
E. Szathmáry (2015):
The divide between prokaryotes and eukaryotes is the biggest known evolutionary discontinuity. There is no space here to enter the whole maze of the recent debate about the origin of the eukaryotic cells; suffice it to say that the picture seems more obscure than 20 y ago. How did eukaryotic life evolve? This is one of the most controversial and puzzling questions in evolutionary history. Life began as single-celled, independent organisms that evolved into cells containing membrane-bound, specialized structures known as organelles. What’s clear is that this new type of cell, the eukaryote, is more complex than its predecessors. What’s unclear is how these changes took place. 24
A.Kauko (2018): The origin of eukaryotes is one of the central transitions in the history of life; without eukaryotes there would be no complex multicellular life.36
F.Rana (2019):
The origin of eukaryotes is one of the hardest and most intriguing problems in the study of the evolution of life, and arguably, in the whole of biology. On average, the volume of eukaryotic cells is about 15,000 times larger than that of prokaryotic cells.30Josip Skejo (2021): Eukaryotic cells
are vastly more complex than prokaryotic cells as evident by their endomembrane system 26
A. Spang (2022): Archaea and Bacteria are often referred to as primary domains of life while eukaryotes form a secondary domain of life. The prevalence of horizontal gene transfer (HGT) via both mobile genetic elements (MGEs) and viruses but also directly between distinct organisms has to some extent questioned the concept of a Tree of Life (TOL), which may be more correctly represented as a network including both vertical and horizontal branches.
Arizona State University (2022): The transition from prokaryote to eukaryote
has remained a central mystery biologists are still trying to untangle. How this crucial transition came to be remains a central mystery in biology.40
Origin of eukaryotesM. A. O’Malley (2015): There are very roughly two main hypotheses for the evolution of eukaryotes: one sees the process as mutation-driven, with lateral acquisitions of genes and organisms also involved but in a causally secondary way; the other sees eukaryogenesis as driven causally by the acquisition of the mitochondrion. The acquisition of the mitochondrion is often portrayed as a one-off event that instigated a rapid transformation with major evolutionary outcomes 38
Eugene V. Koonin (2015): The origin of eukaryotes is
one of the hardest and most intriguing problems in the study of the evolution of life, and arguably, in the whole of biology. Compared to archaea and bacteria (collectively, prokaryotes), eukaryotic cells display a qualitatively higher level of complexity of intracellular organization. Unlike the great majority of prokaryotes, eukaryotic cells possess an extended system of intracellular membranes that includes the eponymous eukaryotic organelle, the nucleus, and fully compartmentalizes the intracellular space. In eukaryotic cells, proteins, nucleic acids and small molecules are distributed by specific trafficking mechanisms rather than by free diffusion as is largely the case in bacteria and archaea. Thus, eukaryotic cells
function on different physical principles compared to prokaryotic cells, which is directly due to their (comparatively) enormous size. The gulf between the cellular organizations of eukaryotes and prokaryotes is all the more striking because
no intermediates have been found. The actin and tubulin cytoskeletons, the nuclear pore, the spliceosome, the proteasome, and the ubiquitin signalling system are only a few of the striking examples of the organizational complexity that seems to be a ‘birthright’ of eukaryotic cells.
The formidable problem that these fundamental complex features present to evolutionary biologists makes Darwin’s famous account of the evolution of the eye look like a simple, straightforward case. Indeed, so intimidating is the challenge of eukaryogenesis that
the infamous notion of ‘irreducible complexity’ has sneaked into serious scientific debate: C. G. Kurland: Genomics and the Irreducible Nature of Eukaryote Cells (2006): Data from many sources give
no direct evidence that eukaryotes evolved by genome fusion between archaea and bacteria. Because their cells appear simpler, prokaryotes have traditionally been considered ancestors of eukaryotes. Here, we review recent data from proteomics and genome sequences
suggesting that eukaryotes are a unique primordial lineage. Mitochondria, mitosomes, and hydrogenosomes are a related family of organelles that distinguish eukaryotes from all prokaryotes. Recent analyses also suggest that early eukaryotes had many introns, and RNAs and proteins found in modern spliceosomes. Nuclei, nucleoli, Golgi apparatus, centrioles, and endoplasmic reticulum are examples of cellular signature structures (CSSs) that distinguish eukaryote cells from archaea and bacteria. Comparative genomics, aided by proteomics of CSSs such as the mitochondria, nucleoli, and spliceosomes, reveals
hundreds of proteins with no orthologs (Orthologs are genes in different species that evolved from a common ancestral gene by speciation)
evident in the genomes of prokaryotes; these are the eukaryotic signature proteins (ESPs). The many ESPs within the subcellular structures of eukaryote cells provide landmarks to track the trajectory of eukaryote genomes from their origins. In contrast, hypotheses that attribute eukaryote origins to genome fusion between archaea and bacteria are
surprisingly uninformative about the emergence of the cellular and genomic signatures of eukaryotes (CSSs and ESPs). The failure of genome fusion to directly explain any characteristic feature of the eukaryote cell is a critical starting point for studying eukaryote origins. 34









