

http://redwood.berkeley.edu/bruno/animal-eyes/Kaas_revised_2013.pdf
Charles Darwin : On the Origin of Species:
Charles Darwin : On the Origin of Species:
To suppose that the eye with all its inimitable contrivances for adjusting the focus to different distances, for admitting different amounts of light, and for the correction of spherical and chromatic aberration, could have been formed by natural selection, seems, I freely confess, absurd in the highest degree. When it was first said that the sun stood still and the world turned round, the common sense of mankind declared the doctrine false; but the old saying of Vox populi, vox Dei, as every philosopher knows, cannot be trusted in science. Reason tells me, that if numerous gradations from a simple and imperfect eye to one complex and perfect can be shown to exist, each grade being useful to its possessor, as is certainly the case; if further, the eye ever varies and the variations be inherited, as is likewise certainly the case; and if such variations should be useful to any animal under changing conditions of life, then the difficulty of believing that a perfect and complex eye could be formed by natural selection, though insuperable by our imagination, should not be considered as subversive of the theory. How a nerve comes to be sensitive to light, hardly concerns us more than how life itself originated; but I may remark that, as some of the lowest organisms, in which nerves cannot be detected, are capable of perceiving light, it does not seem impossible that certain sensitive elements in their sarcode should become aggregated and developed into nerves, endowed with this special sensibility.
How could such a complicated piece of optical machinery arise through a process that has no foresight or intentionality?
When asked, atheists generally point out to Nillson and Penger's Paper from 1994 ,
where the paper makes following assertion :
http://www.rpgroup.caltech.edu/courses/aph161/Handouts/Nilsson1994.pdf
The evolution of complex structures, however, involves modifications of a large number of separate quantitative characters, and in addition there may be discrete innovations and an unknown number of hidden but necessary phenotypic changes. These complications seem effectively to prevent evolution rate estimates for entire organs and other complex structures. An eye is unique in this respect because the structures necessary for image formation, although there may be several, are all typically quantitative in their nature, and can be treated as local modifications of pre-existing tissues. Taking a patch of pigmented light-sensitive epithelium as the starting point, we avoid the more inaccessible problem of photoreceptor cell evolution. Thus, if the objective is limited to finding the number of generations required for the evolution of an eye’s optical geometry, then the problem becomes solvable.
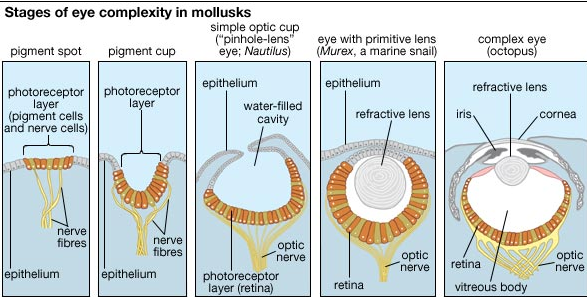
Steps in the evolution of the eye as reflected in the range of eye complexity in living mollusk species (left to right): a pigment spot, as in the limpet Patella; a pigment cup, as in the slit shell mollusk Pleurotomaria; the "pinhole-lens" eye of Nautilus; a primitive lensed eye, as in the marine snail Murex; and the complex eye—with iris, crystalline lens, and retina—of octopuses and squids.
http://skeptoid.com/blog/2013/12/24/is-the-human-eye-irreducibly-complex/
Here’s an abbreviated version of the leading model:
A mutation resulted in a single photoreceptor cell, which allowed the organism to respond to light, and helped to calibrate circadian rhythms by detecting daylight.
Over successive generations, possessing multiple photoreceptors became the norm in the gene pool, because individuals with mutations encoding for an increased number of photoreceptors were better able to react to their surroundings. An arms race began, fueling the evolution of the new sensory organ.
Eventually, what was once just a single photoreceptor cell became a light-sensitive patch. At this point, the creature was still only able to distinguish light from dark.
A slight depression in the patch created a pit, for the first time allowing a limited ability to sense from which direction light or shadow was coming from.
The pit’s opening gradually narrowed to create an aperture — like that of a pinhole camera — making vision sharper.
The aqueous humour formed. A colourless, gelatinous mass filling the chamber of the eye, it helped to maintain the shape of the eye and keep the light sensitive retina in place.
A transparent tissue formed at the front, with a concave curvature for refracting light. The addition of this simple lens drastically improved image fidelity.
A transparent layer evolved in front of the lens. This transparent layer, the cornea, further focused light, and also allowed for more blood vessels, better circulation, and larger eyes.
Behind the cornea, a circular ring formed, the iris, with a hole in its centre, the pupil. By constricting, the iris was able to control the amount of light that reached the retina through the pupil.
Separation of these two layers allowed another gelatinous mass to form, the aqueous humor, which further increased refractive power.
http://www.reviewevolution.com/press/pressRelease_EyeEvolution.php
Since Nilsson and Pelger's article was published, it has been widely--but erroneously--reported that their conclusions were based on a computer model. Berlinski calls this claim "an urban myth." At a minimum, PBS should make clear to viewers that Nilsson's conclusions are not based on computer models at all, and it should acknowledge that his work is highly speculative."
http://creationwiki.org/The_eye_is_too_complex_to_have_evolved_%28Talk.Origins%29
http://www.grisda.org/origins/21039.htm
"What is one to make of all this? First, comparing the evolution of the eye to shape changes on a computer screen seems rather far-fetched. The entire project seems closer to an exercise in geometry than in biology. Second, the exercise assumes a functional starting point. Thus it has nothing to do with the origin of the biochemical systems of vision or the requisite neural network. Third, Nilsson and Pelger's computer exercise operates as if each 1% change in morphology can be accounted for by a single gene mutation. They do not consider the effects of pleiotropy, genetic background, or developmental processes. Fourth, an important part of the model relies on the special circumstance of a layer of clear cells covering the "retina." This layer somehow assumes the proper shape of a lens. Fifth, as noted by the authors, several features of the eye remain unaccounted for, such as the iris. Basically, the only result achieved was to show that two light-sensitive surfaces that differ in shape by 1% will have different efficiencies in photoreception, and that an uninterrupted series of 1% improvements is possible. The failure of scientists to produce new structures in selection experiments illustrates the implausibility of Nilsson and Pelger's "just so" story."
http://www.detectingdesign.com/humaneye.html
If a change in selective pressures favored a dimpled eyespot with a slight increase in visual acuity, pretty soon the majority of the population would have dimpled eyespots. The problem with this notion is that no population of creatures with flat eyespots shows any sort of intra-population range like this were even a small portion of the population has dimpled eyespots to any selectable degree. This is a common assertion, but it just isn't true.
A big problem with these morphologic steps is that they do not take into consideration the fact that vision is more involved than what goes on just within the eye. In order to take any advantage of improved visual acuity within the eye, the brain must also change in such a way that it is able to interpret the information the eye is sending it. Otherwise, if the brain is still step up to appreciate only differences in light from dark sent from the eye, without being able to interpret specific patterns of light and dark on the retina, there would be no selective advantage from a dimpled vs. a flat eyespot. Because of this requirement, whatever evolution happens to take place in the eye, must be backed up by equivalent evolution in brain development and interpretive powers.
the eye/brain is not only a interdependent system:
http://reasonandscience.heavenforum.org/t1638-eye-brain-is-a-interdependent-and-irreducible-complex-system
but the eye is also irreducibly complex in many ways, which can be illustrated best with the Signal transduction pathway in photoreceptor cell's:
http://reasonandscience.heavenforum.org/t1696-photoreceptor-cells
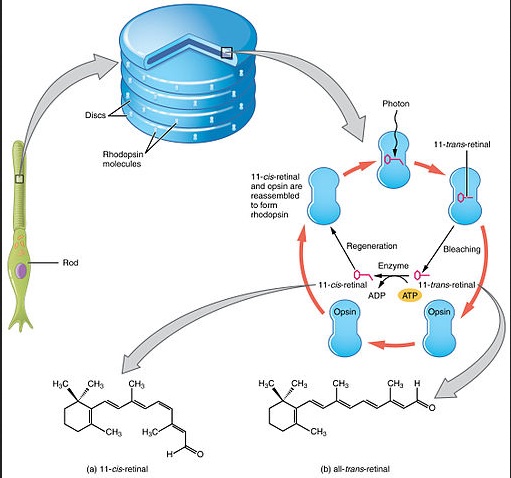
The absorption of light leads to a isomeric change in the retinal molecule.
The signal transduction pathway is the mechanism by which the energy of a photon signals a mechanism in the cell that leads to its electrical polarization. This polarization ultimately leads to either the transmittance or inhibition of a neural signal that will be fed to the brain via the optic nerve. The steps, or signal transduction pathway, in the vertebrate eye's rod and cone photoreceptors are then:
1.The rhodopsin or iodopsin in the disc membrane of the outer segment absorbs a photon, changing the configuration of a retinal Schiff base cofactor inside the protein from the cis-form to the trans-form, causing the retinal to change shape.
2.This results in a series of unstable intermediates, the last of which binds stronger to the G protein in the membrane and activates transducin, a protein inside the cell. This is the first amplification step – each photoactivated rhodopsin triggers activation of about 100 transducins. (The shape change in the opsin activates a G protein called transducin.)
3.Each transducin then activates the enzyme cGMP-specific phosphodiesterase (PDE).
4.PDE then catalyzes the hydrolysis of cGMP to 5' GMP. This is the second amplification step, where a single PDE hydrolyses about 1000 cGMP molecules.
5.The net concentration of intracellular cGMP is reduced (due to its conversion to 5' GMP via PDE), resulting in the closure of cyclic nucleotide-gated Na+ ion channels located in the photoreceptor outer segment membrane.
6.As a result, sodium ions can no longer enter the cell, and the photoreceptor outer segment membrane becomes hyperpolarized, due to the charge inside the membrane becoming more negative.
7.This change in the cell's membrane potential causes voltage-gated calcium channels to close. This leads to a decrease in the influx of calcium ions into the cell and thus the intracellular calcium ion concentration falls.
8.A decrease in the intracellular calcium concentration means that less glutamate is released via calcium-induced exocytosis to the bipolar cell (see below). (The decreased calcium level slows the release of the neurotransmitter glutamate, which can either excite or inhibit the postsynaptic bipolar cells.)
9.Reduction in the release of glutamate means one population of bipolar cells will be depolarized and a separate population of bipolar cells will be hyperpolarized, depending on the nature of receptors (ionotropic or metabotropic) in the postsynaptic terminal (see receptive field).
http://www.detectingdesign.com/humaneye.html
The question now of course is, how could such a system evolve gradually? All the pieces must be in place simultaneously. For example, what good would it be for an earthworm that has no eyes to suddenly evolve the protein 11-cis-retinal in a small group or "spot" of cells on its head? These cells now have the ability to detect photons, but so what? What benefit is that to the earthworm? Now, lets say that somehow these cells develop all the needed proteins to activate an electrical charge across their membranes in response to a photon of light striking them. So what?! What good is it for them to be able to establish an electrical gradient across their membranes if there is no nervous pathway to the worm's minute brain? Now, what if this pathway did happen to suddenly evolve and such a signal could be sent to the worm's brain. So what?! How is the worm going to know what to do with this signal? It will have to learn what this signal means. Learning and interpretation are very complicated processes involving a great many other proteins in other unique systems. Now the earthworm, in one lifetime, must evolve the ability to pass on this ability to interpret vision to its offspring. If it does not pass on this ability, the offspring must learn as well or vision offers no advantage to them. All of these wonderful processes need regulation. No function is beneficial unless it can be regulated (turned off and on). If the light sensitive cells cannot be turned off once they are turned on, vision does not occur. This regulatory ability is also very complicated involving a great many proteins and other molecules - all of which must be in place initially for vision to be beneficial.
http://darwins-god.blogspot.com.br/search?q=eye
When in doubt, doubt the data. Paleontologists agree that the fossil record reveals abrupt appearances, but when convenient evolutionists can always protect their theory with those gaps in the fossil record.
Evolutionary thinking is remarkable. I am reminded of John Earman’s remarks about Hume’s arguments. For it is astonishing how well evolution is treated, given how completely the confection collapses under a little probing. So if Benjamin Radford really is interested in why people believe things for which there is little or no evidence, we have just the topic for him.
How could such a complicated piece of optical machinery arise through a process that has no foresight or intentionality?
When asked, atheists generally point out to Nillson and Penger's Paper from 1994 ,
where the paper makes following assertion :
http://www.rpgroup.caltech.edu/courses/aph161/Handouts/Nilsson1994.pdf
The evolution of complex structures, however, involves modifications of a large number of separate quantitative characters, and in addition there may be discrete innovations and an unknown number of hidden but necessary phenotypic changes. These complications seem effectively to prevent evolution rate estimates for entire organs and other complex structures. An eye is unique in this respect because the structures necessary for image formation, although there may be several, are all typically quantitative in their nature, and can be treated as local modifications of pre-existing tissues. Taking a patch of pigmented light-sensitive epithelium as the starting point, we avoid the more inaccessible problem of photoreceptor cell evolution. Thus, if the objective is limited to finding the number of generations required for the evolution of an eye’s optical geometry, then the problem becomes solvable.
Steps in the evolution of the eye as reflected in the range of eye complexity in living mollusk species (left to right): a pigment spot, as in the limpet Patella; a pigment cup, as in the slit shell mollusk Pleurotomaria; the "pinhole-lens" eye of Nautilus; a primitive lensed eye, as in the marine snail Murex; and the complex eye—with iris, crystalline lens, and retina—of octopuses and squids.
http://skeptoid.com/blog/2013/12/24/is-the-human-eye-irreducibly-complex/
Here’s an abbreviated version of the leading model:
A mutation resulted in a single photoreceptor cell, which allowed the organism to respond to light, and helped to calibrate circadian rhythms by detecting daylight.
Over successive generations, possessing multiple photoreceptors became the norm in the gene pool, because individuals with mutations encoding for an increased number of photoreceptors were better able to react to their surroundings. An arms race began, fueling the evolution of the new sensory organ.
Eventually, what was once just a single photoreceptor cell became a light-sensitive patch. At this point, the creature was still only able to distinguish light from dark.
A slight depression in the patch created a pit, for the first time allowing a limited ability to sense from which direction light or shadow was coming from.
The pit’s opening gradually narrowed to create an aperture — like that of a pinhole camera — making vision sharper.
The aqueous humour formed. A colourless, gelatinous mass filling the chamber of the eye, it helped to maintain the shape of the eye and keep the light sensitive retina in place.
A transparent tissue formed at the front, with a concave curvature for refracting light. The addition of this simple lens drastically improved image fidelity.
A transparent layer evolved in front of the lens. This transparent layer, the cornea, further focused light, and also allowed for more blood vessels, better circulation, and larger eyes.
Behind the cornea, a circular ring formed, the iris, with a hole in its centre, the pupil. By constricting, the iris was able to control the amount of light that reached the retina through the pupil.
Separation of these two layers allowed another gelatinous mass to form, the aqueous humor, which further increased refractive power.
http://www.reviewevolution.com/press/pressRelease_EyeEvolution.php
Since Nilsson and Pelger's article was published, it has been widely--but erroneously--reported that their conclusions were based on a computer model. Berlinski calls this claim "an urban myth." At a minimum, PBS should make clear to viewers that Nilsson's conclusions are not based on computer models at all, and it should acknowledge that his work is highly speculative."
http://creationwiki.org/The_eye_is_too_complex_to_have_evolved_%28Talk.Origins%29
http://www.grisda.org/origins/21039.htm
"What is one to make of all this? First, comparing the evolution of the eye to shape changes on a computer screen seems rather far-fetched. The entire project seems closer to an exercise in geometry than in biology. Second, the exercise assumes a functional starting point. Thus it has nothing to do with the origin of the biochemical systems of vision or the requisite neural network. Third, Nilsson and Pelger's computer exercise operates as if each 1% change in morphology can be accounted for by a single gene mutation. They do not consider the effects of pleiotropy, genetic background, or developmental processes. Fourth, an important part of the model relies on the special circumstance of a layer of clear cells covering the "retina." This layer somehow assumes the proper shape of a lens. Fifth, as noted by the authors, several features of the eye remain unaccounted for, such as the iris. Basically, the only result achieved was to show that two light-sensitive surfaces that differ in shape by 1% will have different efficiencies in photoreception, and that an uninterrupted series of 1% improvements is possible. The failure of scientists to produce new structures in selection experiments illustrates the implausibility of Nilsson and Pelger's "just so" story."
http://www.detectingdesign.com/humaneye.html
If a change in selective pressures favored a dimpled eyespot with a slight increase in visual acuity, pretty soon the majority of the population would have dimpled eyespots. The problem with this notion is that no population of creatures with flat eyespots shows any sort of intra-population range like this were even a small portion of the population has dimpled eyespots to any selectable degree. This is a common assertion, but it just isn't true.
A big problem with these morphologic steps is that they do not take into consideration the fact that vision is more involved than what goes on just within the eye. In order to take any advantage of improved visual acuity within the eye, the brain must also change in such a way that it is able to interpret the information the eye is sending it. Otherwise, if the brain is still step up to appreciate only differences in light from dark sent from the eye, without being able to interpret specific patterns of light and dark on the retina, there would be no selective advantage from a dimpled vs. a flat eyespot. Because of this requirement, whatever evolution happens to take place in the eye, must be backed up by equivalent evolution in brain development and interpretive powers.
the eye/brain is not only a interdependent system:
http://reasonandscience.heavenforum.org/t1638-eye-brain-is-a-interdependent-and-irreducible-complex-system
but the eye is also irreducibly complex in many ways, which can be illustrated best with the Signal transduction pathway in photoreceptor cell's:
http://reasonandscience.heavenforum.org/t1696-photoreceptor-cells
The absorption of light leads to a isomeric change in the retinal molecule.
The signal transduction pathway is the mechanism by which the energy of a photon signals a mechanism in the cell that leads to its electrical polarization. This polarization ultimately leads to either the transmittance or inhibition of a neural signal that will be fed to the brain via the optic nerve. The steps, or signal transduction pathway, in the vertebrate eye's rod and cone photoreceptors are then:
1.The rhodopsin or iodopsin in the disc membrane of the outer segment absorbs a photon, changing the configuration of a retinal Schiff base cofactor inside the protein from the cis-form to the trans-form, causing the retinal to change shape.
2.This results in a series of unstable intermediates, the last of which binds stronger to the G protein in the membrane and activates transducin, a protein inside the cell. This is the first amplification step – each photoactivated rhodopsin triggers activation of about 100 transducins. (The shape change in the opsin activates a G protein called transducin.)
3.Each transducin then activates the enzyme cGMP-specific phosphodiesterase (PDE).
4.PDE then catalyzes the hydrolysis of cGMP to 5' GMP. This is the second amplification step, where a single PDE hydrolyses about 1000 cGMP molecules.
5.The net concentration of intracellular cGMP is reduced (due to its conversion to 5' GMP via PDE), resulting in the closure of cyclic nucleotide-gated Na+ ion channels located in the photoreceptor outer segment membrane.
6.As a result, sodium ions can no longer enter the cell, and the photoreceptor outer segment membrane becomes hyperpolarized, due to the charge inside the membrane becoming more negative.
7.This change in the cell's membrane potential causes voltage-gated calcium channels to close. This leads to a decrease in the influx of calcium ions into the cell and thus the intracellular calcium ion concentration falls.
8.A decrease in the intracellular calcium concentration means that less glutamate is released via calcium-induced exocytosis to the bipolar cell (see below). (The decreased calcium level slows the release of the neurotransmitter glutamate, which can either excite or inhibit the postsynaptic bipolar cells.)
9.Reduction in the release of glutamate means one population of bipolar cells will be depolarized and a separate population of bipolar cells will be hyperpolarized, depending on the nature of receptors (ionotropic or metabotropic) in the postsynaptic terminal (see receptive field).
http://www.detectingdesign.com/humaneye.html
The question now of course is, how could such a system evolve gradually? All the pieces must be in place simultaneously. For example, what good would it be for an earthworm that has no eyes to suddenly evolve the protein 11-cis-retinal in a small group or "spot" of cells on its head? These cells now have the ability to detect photons, but so what? What benefit is that to the earthworm? Now, lets say that somehow these cells develop all the needed proteins to activate an electrical charge across their membranes in response to a photon of light striking them. So what?! What good is it for them to be able to establish an electrical gradient across their membranes if there is no nervous pathway to the worm's minute brain? Now, what if this pathway did happen to suddenly evolve and such a signal could be sent to the worm's brain. So what?! How is the worm going to know what to do with this signal? It will have to learn what this signal means. Learning and interpretation are very complicated processes involving a great many other proteins in other unique systems. Now the earthworm, in one lifetime, must evolve the ability to pass on this ability to interpret vision to its offspring. If it does not pass on this ability, the offspring must learn as well or vision offers no advantage to them. All of these wonderful processes need regulation. No function is beneficial unless it can be regulated (turned off and on). If the light sensitive cells cannot be turned off once they are turned on, vision does not occur. This regulatory ability is also very complicated involving a great many proteins and other molecules - all of which must be in place initially for vision to be beneficial.
http://darwins-god.blogspot.com.br/search?q=eye
When in doubt, doubt the data. Paleontologists agree that the fossil record reveals abrupt appearances, but when convenient evolutionists can always protect their theory with those gaps in the fossil record.
Evolutionary thinking is remarkable. I am reminded of John Earman’s remarks about Hume’s arguments. For it is astonishing how well evolution is treated, given how completely the confection collapses under a little probing. So if Benjamin Radford really is interested in why people believe things for which there is little or no evidence, we have just the topic for him.

