


The possible mechanisms to explain the origin of life
https://reasonandscience.catsboard.com/t2515-abiogenesis-the-possible-mechanisms-to-explain-the-origin-of-life
Either life just coalesced from atomic building blocks through a random fluke collision of disorderly pieces, emerging by “dumb, blind” mechanical processes, a fortuitous accident, spontaneously through self-organization by unguided, non-designed, unintended stochastic coincidence, natural events that turned into self-organization in an orderly manner without external direction, chemical non-biological, purely physico-dynamic kinetic processes and reactions influenced by environmental parameters, or through the direct intervention, direction-giving creative force, and design activity of an intelligent cognitive agency, a powerful conscious creator with intentions, inventive power, will, foreseeing goals and foresight, able to instantiate and create successful solutions in a planned manner.
Rational Wiki: Often brought up in the origins debate is how evolution does not explain the origin of life. Let's get something abundantly clear: abiogenesis and evolution are two completely different things. The theory of evolution says absolutely nothing about the origin of life. It merely describes the processes that take place once life has started.
https://rationalwiki.org/wiki/Abiogenesis
Koonin, E. V. (2012): The emergence of the first replicator system, which represented the “Darwinian breakthrough,” was inevitably preceded by a succession of complex, difficult steps for which biological evolutionary mechanisms were not accessible. The Logic of Chance: The Nature and Origin of Biological Evolution. Amazon.
ADDY PROSS (2012): Darwinian theory is a biological theory and therefore deals with biological systems, whereas the origin of life problem is a chemical problem, and chemical problems are best solved with chemical (and physical) theories. Attempting to explain chemical phenomena with biological concepts is methodologically problematic.
What is Life?: How Chemistry Becomes Biology
Paul Davies (2021): I think in all honesty a lot of people even confuse it the people who aren't familiar with the area that oh I presume Darwinian evolution sort of accounts for the origin of life but of course, you don't get an evolutionary process until you've got a self-replicating molecule. ( Darwin ) gave us a theory of evolution about how life has evolved but he uh didn't want to tangle with how you go from non-life to life and for me, that's a much bigger step. Why Darwinian evolution does NOT explain the origin of life
Fry, Iris. (2010): The Role of Natural Selection in the Origin of Life. Origins of Life and Evolution of Biospheres Link.
Iris Fry analyzes various theories on the origin of life, including RNA-first, metabolism-first, and others. She concludes that while none of these paradigms have decisive experimental support, gene-first theories show potential. As of her writing, no functioning system of genetic replication had been achieved without the addition of an external protein enzyme.
Alan W. Schwartz (2007): A problem that is familiar to organic chemists is the production of unwanted byproducts in synthetic reactions. For prebiotic chemistry, where the goal is often the simulation of conditions on the prebiotic Earth and the modeling of a spontaneous reaction, it is not surprising – but nevertheless frustrating – that the unwanted products may consume most of the starting material and lead to nothing more than an intractable mixture, or -gunk.. The most well-known examples of the phenomenon can be summarized quickly: Although the Miller –Urey reaction produces an impressive set of amino acids and other biologically significant compounds, a large fraction of the starting material goes into a brown, tar-like residue that remains uncharacterized; i.e., gunk. While 15% of the carbon can be traced to specific organic molecules, the rest seems to be largely intractable Even if we focus only on the soluble products, we still have to deal with an extremely complex mixture of compounds. The carbonaceous chondrites, which represent an alternative source of starting material for prebiotic chemistry on Earth, and must have added enormous quantities of organic material to the Earth at the end of the Late Heavy Bombardment (LHB), do not offer a solution to the problem just referred to. The organic material present in carbonaceous meteorites is a mixture of such complexity that much ingenuity has gone into the design of suitable extraction methods, to isolate the most important classes of soluble (or solubilized) components for analysis. Whatever the exact nature of an RNA precursor which may have become the first selfreplicating molecule, how could the chemical homogeneity which seems necessary to permit this kind of mechanism to even come into existence have been achieved? What mechanism would have selected for the incorporation of only threose, or ribose, or any particular building block, into short oligomers which might later have undergone chemically selective oligomerization? Virtually all model prebiotic syntheses produce mixtures. 2
A.G.Cairns-Smith (1985): It is one of the most singular features of the unity of biochemistry that this mere convention is universal. Where did such agreement come from? You see non-biological processes do not as a rule show any bias one way or the other, and it has proved particularly difficult to see any realistic way in which any of the constituents of a 'prebiotic soup' would have had predominantly 'left-handed' or right-handed' molecules. It is thus particularly difficult to see this feature as having been imposed by initial conditions. 3
A.G.Cairns-Smith (1985): genetic takeover, page 70: Suppose that by chance some particular coacervate droplet in a primordial ocean happened to have a set of catalysts, etc. that could convert carbon dioxide into D-glucose. Would this have been a major step forward towards life? Probably not. Sooner or later the droplet would have sunk to the bottom of the ocean and never have been heard of again. It would not have mattered how ingenious or life-like some early system was; if it lacked the ability to pass on to offspring the secret of its success then it might as well never have existed. So I do not see life as emerging as a matter of course from the general evolution of the cosmos, via chemical evolution, in one grand gradual process of complexification. Instead, following Muller (1929) and others, I would take a genetic View and see the origin of life as hinging on a rather precise technical puzzle. What would have been the easiest way that hereditary machinery could have formed on primitive Earth? 4
PROFESSOR DR KLAUS DOSE: The Origin of Life: More Questions Than Answers
Evolution, as used here, refers to any development or change influenced by the environment. The term "chemical evolution" specifically pertains to changes in chemical substances, indicating that fundamental transformations occur within molecules. Often, it is employed as a synonym for the "abiotic" or "prebiotic formation" of organic molecules in cosmic systems, particularly in the context of the early Earth. It is assumed that these organic molecules were produced from the constituents present in the primitive atmosphere, hydrosphere, and, to some extent, the lithosphere. On the other hand, "molecular evolution" encompasses a broader scope than chemical evolution. It encompasses self-assembly into more complex structures, such as membranes, protocells, cell-like systems, and protocellular organelles. Additionally, it involves the subsequent evolution of proto-cells or protobionts, leading to the emergence of the first modern cells, also known as "Urzellen" or progenotes. The transition from protobionts to progenotes has been referred to as proto-Darwinian evolution. In contrast, "Darwinian evolution" pertains to the evolution from progenotes to the vast array of contemporary cells and organisms.
https://sci-hub.ee/10.1179/isr.1988.13.4.348
Comment: According to the description provided, "chemical evolution" primarily focuses on the changes in chemical substances and the fundamental transformations that occur within molecules. It is often used as a term synonymous with the "abiotic" or "prebiotic formation" of organic molecules in cosmic systems, particularly in the context of the early Earth. The key aspect of chemical evolution is the formation of organic molecules from inorganic precursors found in the primitive atmosphere, hydrosphere, and lithosphere. Notably, the concept of "chemical evolution" does not explicitly include natural selection, which is a fundamental mechanism of Darwinian evolution. Natural selection is a process that acts on existing life forms, favoring certain traits or characteristics that provide a reproductive advantage in a given environment. Over time, this leads to the accumulation of beneficial traits within a population, leading to the adaptation and diversification of organisms. While chemical evolution sets the stage by forming the necessary building blocks of life, such as amino acids and nucleotides, it does not inherently involve the process of natural selection. Instead, it provides the foundation upon which subsequent evolutionary processes, like "molecular evolution" and "Darwinian evolution," can take place. "Molecular evolution," as mentioned in the initial description, goes beyond chemical evolution by encompassing the self-assembly of complex structures and the subsequent evolution of proto-cells to modern cells. This process involves changes in the molecular and structural organization of early life forms, leading to the emergence of more advanced cellular systems. On the other hand, "Darwinian evolution" encompasses the evolution of progenotes (early simple cells) to the vast array of contemporary cells and organisms we observe today. Natural selection is a central driving force in this form of evolution, shaping the diversity of life through the differential survival and reproduction of individuals with advantageous traits.
Self-organization refers to the supposed and hypothesized spontaneous emergence of order and complexity without external direction or control and without the need for external instructions. While self-organization is a hypothesized concept, it has never been directly observed or reproduced in a laboratory setting.
Gennady Shkliarevsky: THE UNIVERSAL EVOLUTION AND THE ORIGIN OF LIFE 2021
Most current OOL perspectives invoke chance or coincidence in their explanations of the origin of life. Sean Carroll, a well-known evolutionary biologist, refers to the emergence of life as “the mother of all accidents” and “the accident of all mothers.” There are also additional problems that plague the dominant scenario on the origin of prokaryotic cells. Most, if not all of them, invoke chance or coincidence to explain the origin of major cell components. Not only that, but they also invoke chance or coincidence to explain why these components came together to form a cell. As has been explained earlier, invoking chance even once is highly problematic; invoking it twice to explain the same phenomenon makes an explanation very questionable. As has been repeatedly pointed out, the liberal use of chance and coincidence as an explanatory mode is also a source of concern in theories about the origin and evolution of early life. In order to explain the emergence of radical novelty, many current OOL perspectives invoke chance or coincidence.
https://arxiv.org/ftp/arxiv/papers/2104/2104.08076.pdf
Alexei A. Sharov: Coenzyme world model of the origin of life 2016 Mar 9
The probability of transferring the full set of coding molecules to descendants by pure chance may be problematic especially if droplets carry too many kinds of coding molecules and some of them are present in a small number of copies. This combinatorial problem can be partially meliorated by the “stochastic corrector” mechanism, which is a preferential propagation of systems with a full set of coding molecules (Szathmáry, 1999). Systems with an incomplete set of coding molecules are more likely to fail in surviving and reproduction because some of their functions appear missing. This kind of stochastic correction is a primordial version of the purifying selection; and like the purifying selection, it reduces the overall reproduction rate of the population.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4875852/
Horst Rauchfuss Chemical Evolution and the Origin of Life
An important, but as yet unsolved, problem is provided by the chirality of the amino acids. The main question is whether the enantiomeric selection of the amino acids took place before or after the evolution of life.
Others see the genetic code as being purely fortuitous, a system that was “frozen” at some time in history.
William Dembski (2002): The problem is that nature has too many options and without design couldn’t sort through all those options. The problem is that natural mechanisms are too unspecific to determine any particular outcome. Natural processes could theoretically form a protein, but also compatible with the formation of a plethora of other molecular assemblages, most of which have no biological significance. Nature allows them full freedom of arrangement. Yet it’s precisely that freedom that makes nature unable to account for specified outcomes of small probability. Nature, in this case, rather than being intent on doing only one thing, is open to doing any number of things. Yet when one of those things is a highly improbable specified event, design becomes the more compelling, better inference. Occam's razor also boils down to an argument from ignorance: in the absence of better information, you use a heuristic to accept one hypothesis over the other. 5
There's nothing about inert chemicals and physical forces that say we want to get life at the end of the abiogenesis process. Molecules do not have the "drive", they do not "want" to find ways to harness the energy and become more efficient
One cannot explain the origin of evolution, through evolution. It is widespread and very common to see the attempt to smuggle the Darwinian dynamic of replication with a heritable variation into the origin of life. Biological evolution by natural selection does and cannot explain the origin of life. Natural selection only acts on the random variation of alleles based on DNA replication, but the origin of genes, and replication is among the origin of the entire self-replicating cell, what the origin of life research has to explain.
1. When we see complexification, that is: Interconnecting parts, weaving together, aggregating subunits, wrapping around, encompassing, interlinking, interlocking, twisting, interlacing, fusing, assembling related things, intricately combining things, where the system is greater than the sum of their parts. then it is logical to attribute such actions to an intelligently acting mind with foresight and foreknowledge, and distant goals.
2. Making systems with the hallmark of complexity depends on the careful elaboration and design in detail of many elementary parts and interconnecting them in a meaningful way conferring a specific purpose or function. Not rarely, small changes in one part of the system can cause sudden and unexpected outputs in other parts of the system, system-wide reorganization, or breaking down of the higher function.
3. Why WOULD a molecule self-replicate apart from duplication performed by the DNA replication machinery? There was no prebiotic natural selection with the goal to survive and reproduction. Why should be assumed that molecules - that are randomly assembled by chemical/physical forces/happenstance and just happened to stay in place - have the drive or goal to self-replicate, to become the complex macromolecular building blocks of living cells? Or to have any goal at all? Why would/should molecules strive toward increasing complexity without any possible "psychological" basis for doing so? Random accidents are not the best case-adequate explanation for the origin of emerging properties of a complex system. Intelligent design is.
Exploring the Deep Mystery of Life's Origins Aug 8, 2022
Nick Lane: Our kind of cell arose once in four billion years of evolution. And it seems to have been something of a bit of a freak accident.
https://www.youtube.com/watch?v=ATubwpnVLAY
Wilhelm T. S. Huck Robustness, Entrainment, and Hybridization in Dissipative Molecular Networks, and the Origin of Life May 30, 2019
Life emerged spontaneously from the selfassembly, or spontaneous organization, of the organic products of reactions, occurring in complex mixtures of molecules formed abiotically from simple precursors and sequences of reactions.
https://robobees.seas.harvard.edu/files/gmwgroup/files/1320.pdf
Phillip E. Johnson, DARWIN ON TRIAL: Darwin persuades us that the seemingly purposeful construction of living things can very often, and perhaps always, be attributed to the operation of natural selection.
If you have things that are reproducing their kind;
if there are sometimes random variations, nevertheless, in the offspring;
if such variations can be inherited;
if some such variations can sometimes confer an advantage on their owners;
if there is competition between the reproducing entities;-
if there is an overproduction so that not all will be able to produce offspring themselves-
then these entities will get better at reproducing their kind. What is needed for natural selection are things that conform to those 'ifs'. Self-replicating cells are prerequisites for evolution. None of this was available prebiotically to explain the origin of the first life form. 1
MARIO VANEECHOUTTE The scientific origin of life 2000
We hypothesize that the origin of life, that is, the origin of the first cell, cannot be explained by natural selection among self-replicating molecules, as is done by the RNA-world hypothesis.
The hypothesis espoused here states that it is virtually impossible that the highly complicated system cell developed gradually around simple self-replicating molecules (RNA-hypercycles or autocatalytic peptide networks) by means of natural selection; as is proposed by, for example, the RNA-world hypothesis. Despite searching quadrillions of molecules, it is clear that a spontaneous RNAreplicator is unlikely to occur. Reports of nucleotide and peptide self-replication still depend upon human intervention (for instance, by changing the environmental conditions between two rounds of replication or by denaturing the double strands). The problem of denaturing the double-nucleotide strand in a nonenzymatic manner has been overlooked and has contributed to a failure to establish molecular self-replication. The first cell, life, was born and natural selection (selection among variations on the theme of autonomous duplication) commenced.
The hypothesis suggested here states that no such autonomous duplication existed before the first cell and, thus, natural selection started only with the first cell.
https://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.645.3011&rep=rep1&type=pdf
Jonathan Sarfati Natural selection cannot explain the origin of life 31 March 2021
Natural selection requires pre-existing life.
https://creation.com/ns-origin-of-life
The first life could emerge by chance in a depression or tide pool, as one of countless reaction systems.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4810237/#:~:text=The%20bottom%2Dup%20approach%20is,%2C%20genetic%20code%2C%20and%20proteins.
Evolution by mutations and natural selection do not explain the origin of life, since evolution depends on the Darwinian dynamic of replication with a heritable variation. The only alternative to design are random unguided lucky events.
Or through the direct intervention and creative force of an intelligent agency, a powerful creator.
In an attempt to explain the origin of life, scientists propose a two-stage process of natural chemical evolution: formation of organic molecules, which combine to make larger biomolecules; self-organization of these molecules into a living organism. The origin of life can not be explained through biological nor chemical evolution. Adaptation, mutation, and natural selection depend on DNA replication. Heredity is guaranteed by faithful DNA replication whereas evolution depends upon errors accompanying DNA replication. Neither can it be explained through physical laws. Life depends on codes and instructional complex information. This information can only be generated by when the arrangement of the code is free and unconstrained, and any of the four bases of the genetic code can be placed in any of the positions in the sequence to generate the information. The only alternative, if the action of a creative agency is excluded, would be spontaneous self-assembly by orderly aggregation of prebiotic elements and building blocks in a sequentially correct manner without external direction.
Origin of life: A prebiotic route to DNA
The first polymeric molecules capable of storing information and reproducing themselves were randomly assembled from organic precursors that were available on the early Earth.
https://www.sciencedaily.com/releases/2019/06/190618103721.htm
Albert L. Lehninger described how inanimate matter crossed the great divide to become alive. “We now come to the critical moment in evolution,” he wrote in 1970, “in which the first semblance of ‘life’ appeared, through the chance association of a number of abiotically formed macromolecular components.”
Natural selection is not a possible mechanism to explain the origin of life, since evolution depends on DNA replication
Life's Origins: What Was the Origin Of Complex Organic Molecules? We are going to make an assumption: the chemical components found in all living prokaryotic and eukaryotic cells was present in the ancestral organism from which all life derived. In other words: the spontaneous interaction of the molecules present in the early earth's atmosphere formed more complex organic molecules, such as amino acids, sugars, fatty acids, and nitrogen bases - the building blocks of life as we know it today.
https://www2.gwu.edu/~darwin/BiSc151/Origin/origin.html
Question : Was there natural selection prior life starting?
Reply: No, natural selection as a mechanism of evolution cannot exist prior to the existence of life. Natural selection is a process by which certain traits that are better suited to an organism's environment become more prevalent in a population over time. This process depends on the existence of organisms that can reproduce, pass on traits to their offspring, and have variations in traits that can be selected for or against based on their survival and reproductive success.
In order for natural selection to occur, there must first be life, and this is where the question of the origin of life becomes relevant. While natural selection can explain the evolution of species once life has arisen, it cannot explain the initial emergence of life from non-living matter.
Stephen C. Meyer, The return of the God hypothesis
The process of natural selection presupposes the differential reproduction of already living organisms and thus a preexisting mechanism of self-replication. Yet self-replication in all extant cells depends upon functional (and therefore sequence-specific, information-rich) proteins and nucleic acids. And the origin of such information-rich molecules is precisely what Oparin needed to explain. Thus, many rejected his postulation of prebiotic natural selection as question-begging. As the evolutionary biologist Theodosius Dobzhansky insisted, “Pre-biological natural selection is a contradiction in terms.” Or as Christian de Duve explained, theories of prebiotic natural selection “need information which implies they have to presuppose what is to be explained in the first place.”
The Origin of the First Hereditary Replicators
This process is still an unsolved problem. By itself, this transition is not an evolutionary one because, without hereditary replicators, no Darwinian evolution is possible.
https://www.pnas.org/content/112/33/10104
Alemi Mario: The Amazing Journey of Reason from DNA to Artificial Intelligence 2020
Darwin probably didn’t propose a theory for the origin of life simply because applying Darwin’s mechanism of natural selection to the emergence of life, as done by Dawkins (1976), is like comparing apples with pears (Johnson 2010). What’s more, the idea that a self-replicating molecule with an information content casually appeared in a primordial soup, as imagined by Dawkins (1976) (“At some point a particularly remarkable molecule was formed by accident. We will call it the Replicator.”) appears to be statistically groundless (Yockey 1977).
https://link.springer.com/book/10.1007%2F978-3-030-25962-4
Is life a gamble? Scientist models universe to find out April 21, 2020
Scientists suspect that the complex life that slithers and crawls through every nook and cranny on Earth emerged from a random shuffling of non-living matter that ultimately spit out the building blocks of life.
https://www.livescience.com/origin-of-life-rna-universe-model.html
A. G. CAIRNS-SMITH Seven clues to the origin of life, page 36:
And if you ask me how the next stage happened, how the smallish 'molecules of life' came together to make the first reproducing evolving being, I will reply: 'With time, and more time, and the resource of oceans.' I will sweep my arms grandly about. 'Because you see. in the absence of oxygen the oceans would have accumulated "the molecules of life". The oceans would have been vast bowls of nutritious soup. Chance could do the rest.
The role of natural selection in the origin of life
Unlike living systems that are products of and participants in evolution, these prebiotic chemical structures were not products of evolution. Not being yet intricately organized, they could have emerged as a result of ordinary physical and chemical processes.
https://www.ncbi.nlm.nih.gov/pubmed/20407927
Alternative Pathways of Carbon Dioxide Fixation: Insights into the Early Evolution of Life? July 6, 2011
The fixation of inorganic carbon into organic material (autotrophy) is a prerequisite for life and sets the starting point of biological evolution.
https://sci-hub.ren/https://www.annualreviews.org/doi/10.1146/annurev-micro-090110-102801
Jack W. Szostak Functional proteins from a random-sequence library
Functional primordial proteins presumably originated from random sequences
https://molbio.mgh.harvard.edu/szostakweb/publications/Szostak_pdfs/Keefe_Szostak_Nature_01.pdf?fbclid=IwAR0giOg_aZfFRKQALk7CB22nVIx32ShiN0Vp78cwtAYwmwQ_0RJicfxpR1M
LIFE The Science of Biology, TENTH EDITION, page 3
When we consider how life might have arisen from nonliving matter, we must take into account the properties of the young Earth’s atmosphere, oceans, and climate, all of which were very different than they are today. Biologists postulate that complex biological molecules first arose through the random physical association of chemicals in that environment.
Neither Evolution nor physical necessity are a driving force prior dna replication. The only two alternatives are either a) creation by an intelligent agency, or b) Random, unguided, undirected natural events by a lucky "accident".
Koonin, the logic of chance, page 246
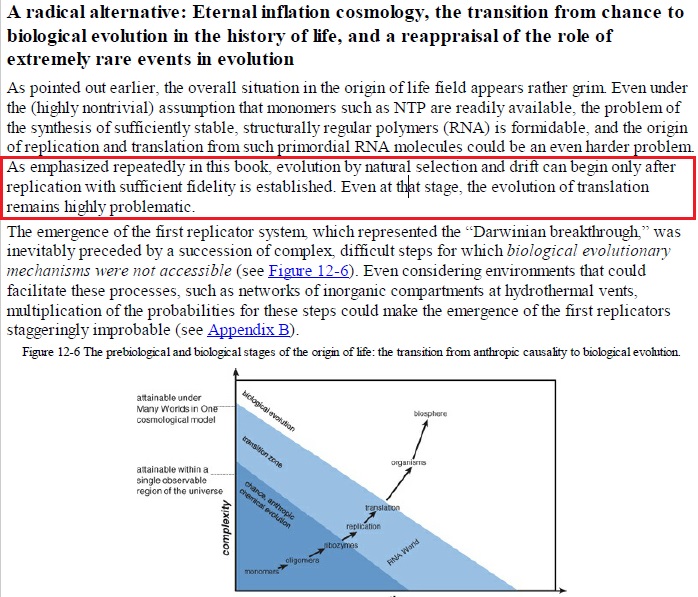
Evolution by natural selection and drift can begin only after replication with sufficient fidelity is established. Even at that stage, the evolution of translation remains highly problematic. The emergence of the first replicator system, which represented the “Darwinian breakthrough,” was inevitably preceded by a succession of complex, difficult steps for which biological evolutionary mechanisms were not accessible . The synthesis of nucleotides and (at least) moderate-sized polynucleotides could not have evolved biologically and must have emerged abiogenically—that is, effectively by chance abetted by chemical selection, such as the preferential survival of stable RNA species. Translation is thought to have evolved later via an ad hoc selective process. Did you read this ???!! An ad-hoc process ??
Without code there can be no self-replication. Without self-replication, you can’t have reproduction. Without reproduction, you can’t have evolution or natural selection.
Heredity is guaranteed by faithful DNA replication whereas evolution depends upon errors accompanying DNA replication. ( Furusawa, 1998 ) We hypothesize that the origin of life, that is, the origin of the first cell, cannot be explained by natural selection among self-replicating molecules, as is done by the RNA-world hypothesis. ( Vaneechoutte M )
Chance and necessity do not explain the origin of life
https://www.academia.edu/1204161/Trevors_J.T._Abel_D.L._2004_Chance_and_necessity_do_not_explain_the_origin_of_life_Cell_Biology_International_28_729-739
Selection pressure cannot select nucleotides at the digital programming level where primary structures form. Genomes predetermine the phenotypes which natural selection only secondarily favors. Contentions that offer nothing more than long periods of time offer no mechanism of explanation for the derivation of genetic programming. No new informationis provided by such tautologies. The argument simply says it happened. As such, it is nothing more than blind belief. Science must provide rational theoretical mechanism, empirical support, prediction fulfillment, or some combination of these three. If none of these three are available, science should reconsider that molecular evolution of genetic cybernetics is a proven fact and press forward with new research approaches which are not obvious at this time. 5
I would like to plead with you, simply, please realize you cannot use the words `natural selection' loosely. Prebiological natural selection is a contradiction of terms."
(Dobzhansky, T.G., Discussion of "Synthesis of Nucleosides and Polynucleotides with Metaphoric Esters,", Oct. 27-30, 1963, Academic Press: New York NY, 1965, pp.309-310).
B.Alberts, Molecular Biology of the Cell, 5th edition, page 406
Self-Replicating Molecules Undergo Natural Selection
The three-dimensional folded structure of a polynucleotide affects its stability, its actions on other molecules, and its ability to replicate. Therefore, certain polynucleotides will be especially successful in any self-replicating mixture. Because errors inevitably occur in any copying process, new variant sequences of these polynucleotides will be generated over time.

Stephen Meyer, Darwin's doubt, page 6:
Natural selection assumes the existence of living organisms with a capacity to reproduce. Yet self-replication in all extant cells depends upon information-rich proteins and nucleic acids (DNA and RNA), and the origin of such information-rich molecules is precisely what origin-of-life research needs to explain. That’s why Theodosius Dobzhansky, one of the founders of the modern neo-Darwinian synthesis, can state flatly, “Pre-biological natural selection is a contradiction in terms.”5 Or, as Nobel Prize–winning molecular biologist and origin-of-life researcher Christian de Duve explains, theories of prebiotic natural selection fail because they “need information which implies they have to presuppose what is to be explained in the first place.
That means, evolution was not a driving force and acting for the emergence and origin of the first living organisms. The only remaining possible mechanisms are chemical reactions acting upon unregulated, aleatory events ( luck, chance), or physical necessity. ( where chemical reactions are forced into taking a certain course of action. )
Morowitz: THE ORIGIN AND NATURE OF LIFE ON EARTH page 18
The Darwinian framework for selection requires support from other error-correcting mechanisms that operate in simpler contexts, to arrive at a mechanism sufficient to explain the emergence, overall organization, and long-term persistence of life from non-living precursors.
In a relatively short time, the ocean became a broth of these molecules, and given enough time, the right combination of molecules came together by pure chance to form a replicating entity of some kind that evolved into modern life.
https://www.americanscientist.org/article/the-origin-of-life
Physical necessity & Physical laws
Stephen C.Meyer, The return of the God hypothesis, page 216:
Rather than having a genetic molecule capable of unlimited novelty, with all the unpredictable and aperiodic sequences that characterize informative texts, we would have a highly repetitive text awash in redundant sequences—much as happens in crystals. Indeed, in a crystal the forces of mutual chemical attraction do completely explain the sequential ordering of the constituent parts. Consequently, crystals cannot convey novel information. Bonding affinities, to the extent they exist, cannot be used to explain the origin of information. Self-organizing chemical affinities generate highly repetitive “order,” but not information; they create mantras, not messages
The nucleotide sequence of DNA and RNA have an instructional function to make proteins and is NOT random but complex and specified, and not due to physical necessity or physical laws. And this is what events in a prebiotic land would need to produce: a minimal set of proteins .... and this kind of specification does not arise through chemical reactions ...... the result of a chemical reaction is not random. But the events dealing with an eventual chemical reaction would have been if there was not a mind guiding the events.
Consider, for example, what would happen if the individual nucleotide bases (A, C, G, T) in the DNA molecule did interact by chemical necessity (along the information-bearing axis of DNA). Suppose that every time adenine (A) occurred in a growing genetic sequence, it attracted cytosine (C) to it,26 which attracted guanine (G), which attracted thymine (T), which attracted adenine (A), and so on. If this were the case, the longitudinal axis of DNA would be peppered with repetitive sequences of ACGT. Rather than being a genetic molecule capable of virtually unlimited novelty and characterized by unpredictable and aperiodic sequences, DNA would contain sequences awash in repetition or redundancy—much like the arrangement of atoms in crystals.
Michael Polanyi: Life's Irreducible Structure,
Science mag, 1968
In Galileo's experiments on balls rolling down a slope, the angle of the slope was not derived from the laws of mechanics, but was chosen by Galileo. And as this choice of slopes was extraneous to the laws of mechanics, so is the shape and manufacture of test tubes extraneous to the laws of chemistry. The same thing holds for machinelike boundaries; their structure cannot be defined in terms of the laws which they harness. Nor can a vocabulary determine the content of a text, and so on. Therefore, if the structure of living things is a set of boundary conditions, this structure is extraneous to the laws of physics and chemistry which the organism is harnessing. Thus the morphology of living things transcends the laws of physics and chemistry.the codelike structure of DNA must be assumed to have come about by a sequence of chance variations established by natural selection. But this evolutionary aspect is irrelevant here; whatever may be the origin of a DNA configuration, it can function as a code only if its order is not due to the forces of potential energy. It must be as physically indeterminate as the sequence of words is on a printed page. As the arrangement of a printed page is extraneous to the chemistry of the printed page, so is the base sequence in a DNA molecule extraneous to the chemical forces at work in the DNA molecule. It is this physical indeterminacy of the sequence that produces the improbability of occurrence of any particular sequence and thereby enables it to have a meaning-a meaning that has a mathematically determinate information content equal to the numerical improbability of the arrangement.
A deterministic answer assumes that the laws of physics and chemistry have causally and sequentially determined the obligatory series of events leading from inanimate matter to life – that each step is causally linked to the previous one and to the next one by the laws of nature. In principle, in a strictly deterministic situation, the state of a system at any point in time determines the future behavior of the system – with no random influences. To invoke a guided determinism toward the formation of life would only make sense if the construction of life was demonstrably a preferential, highly probable natural pathway.
Luisi, The Emergence of Life; From Chemical Origins to Synthetic Biology, page 21
Just like computer codes, the genetic code is arbitrary. There is no law of physics that says “1” has to mean “on” and “0” has to mean “off.” There’s no law of physics that says 10000001 has to code for the letter “A.” Similarly, there is no law of physics that says three Guanine molecules in a row have to code for Glycine. In both cases, the communication system operates from a freely chosen, fixed set of rules.
In all communication systems it is possible to label the encoder, the message and the decoder and determine the rules of the code.
The rules of communication systems are defined in advance by conscious minds. There are no known exceptions to this. Therefore we have 100% inference that the Genetic Code was designed by a conscious mind.
Physical laws which result in physical constraints, where chemical reactions are forced into taking a certain course of action is an often cited possible mechanism for the origin of life.
We are moving from chemistry to biology. Henceforward, life, it goes without saying, is independent of its chemical substrate, and its evolution does not follow paths that are predictable solely based on the laws of physics.
M. Gargaud · H. Martin · P. López-García T. Montmerle · R. Pascal Young Sun, Early Earth and the Origins of Life, page 95
Laurent Boiteau Prebiotic Chemistry: From Simple Amphiphiles to Protocell Models, page 3:
Spontaneous self-assembly occurs when certain compounds associate through noncovalent hydrogen bonds, electrostatic forces, and nonpolar interactions that stabilize orderly arrangements of small and large molecules. The argument that chemical reactions in a primordial soup would not act upon pure chance, and that chemistry is not a matter of "random chance and coincidence, finds its refutation by the fact that the information stored in DNA is not constrained by chemistry. Yockey shows that the rules of any communication system are not derivable from the laws of physics. He continues: “there is nothing in the physicochemical world that remotely resembles reactions being determined by a sequence and codes between sequences.” In other words, nothing in nonliving physics or chemistry obeys symbolic instructions.
Ulrich E. Stegmann: The arbitrariness of the genetic code March 2004 5
Some of the processes expected to involve semantic information are certainly not chemically arbitrary and, therefore, chemical arbitrariness is not a necessary condition for a semantic relation.
The problem of information to explain the origin of life
Norbert Weiner - MIT Mathematician - Father of Cybernetics
"Information is information, not matter or energy. No materialism which does not admit this can survive at the present day."
It has to be explained:
- a library index and fully automated information classification, storage and retrieval program ( chromosomes, and the gene regulatory network )
- The origin of the complex, codified, specified, instructional information stored in the genome and epigenetic codes to make the first living organism
- The origin of the genetic Code
- How it got nearly optimal for allowing additional information within protein-coding sequences
- How it got more robust than 1 million alternative possible codes
- The origin of the over twentythree epigenetic codes
- The origin of the information transmission system, that is the origin of the genetic code itself, encoding, transmission, decoding and translation
- The origin of the genetic cipher/translation, from digital ( DNA / mRNA ) to analog ( Protein )
- The origin of the hardware, that is DNA, RNA, amino acids, and carbohydrates for fuel generation
- The origin of the replication/duplication of the DNA
- The origin of the signal recognition particle
- The origin of the tubulin Code for correct direction to the final destination of proteins
Information theory cannot normally be used to predict how chemicals will react because some chemicals react with each other readily, and others only react very slowly. Others do not react with each other at all. Thus, the likelihood of two chemicals joining together depends on both the quantity of the chemicals present and their chemical properties. Information theory can easily deal with the effects of quantity, but it has no way to deal with chemical properties.
Stuart Pullen, Intelligent Design or Evolution? Why the Origin of Life and the Evolution of Molecular Knowledge Imply Design, page 88
http://lifesorigin.com/prebiotic-evolution4.pdf
Stephen C. Meyer observed:
“There are neither bonds nor bonding affinities—differing in strength or otherwise—that can explain the origin of the base sequencing that constitutes the information in the DNA molecule”
(Signature in the Cell, 243).
As Paul Davies lamented,
“We are still left with the mystery of where biological information comes from.… If the normal laws of physics can’t inject information, and if we are ruling out miracles, then how can life be predetermined and inevitable rather than a freak accident? How is it possible to generate random complexity and specificity together in a lawlike manner? We always come back to that basic paradox”
(Fifth Miracle, 258).
A law of nature could not alone explain how life began, because no conceivable law would compel a legion of atoms to follow precisely a prescribed sequence of assemblage.
Paul Davies, The origin of Life, page 17
Werner Gitt summarized it this way:
“A necessary requirement for generating meaningful information is the ability to select from alternatives and this requires an intelligent, volitional entity.… Unguided, random processes cannot do this—not in any amount of time because this selection process demands continuous guidance by intelligent beings that have a purpose”
(Without Excuse, 50–51).
Let's assume that we begin with the sequence R-T-X, and will add two amino acids "B" and "A" to it. If amino acid "B" is the most reactive amino acid, the sequence would be R-T-X-B-A. However, if "A" is the most reactive amino acid, then the sequence would be R-T-X-A-B. In a random chemical reaction, the sequence of amino acids would be determined by the relative reactivity of the different amino acids. The polymer chain found in natural proteins and DNA has a very precise sequence that does not correlate with the individual components' reaction rates. Since all of the amino acids have relatively similar structures, they all have similar reaction rates; they will all react at about the same rate making the precise sequence by random chemical reactions unthinkably unlikely. This is the problem of Chemical Reactivity.
http://www.icr.org/article/evolution-hopes-you-dont-know-chemistry-problem-co/
The Genetic Code
http://hyperphysics.phy-astr.gsu.edu/hbase/Organic/gencode.html
DNA contains a true code. Being a true code means that the code is free and unconstrained; any of the four bases can be placed in any of the positions in the sequence of bases. Their sequence is not determined by the chemical bonding. There are hydrogen bonds between the base pairs and each base is bonded to the sugar phosphate backbone, but there are no bonds along the longitudional axis of DNA. The bases occur in the complementary base pairs A-T and G-C, but along the sequence on one side the bases can occur in any order, like the letters of a language used to compose words and sentences. Since nucleotides can be arranged freely into any informational sequence, physical necessity could not be a driving mechanism.
Abiogenesis is the process by which life arises naturally from non-living matter. Scientists speculate that life may have arisen as a result of random chemical processes happening to produce self-replicating molecules.
http://rationalwiki.org/wiki/Abiogenesis
Paul Davies conceded, “Unfortunately, before Darwinian evolution can start, a certain minimum level of complexity is required. But how was this initial complexity achieved? When pressed, most scientists wring their hands and mutter the incantation ‘Chance.’ So, did chance alone create the first self-replicating molecule?” (Fifth Miracle, 138).
If design or physical necessity is discarded, the only remaining possible mechanism for the origin of life is chance/luck.
Emergent properties
Claim: The origin of life could be an emergent property of basic chemical reactions
Reply: An emergent property is a property that a collection or complex system has, but which the individual members do not have. In biology, for example, heart is made of heart cells, heart cells on their own don't have the property of pumping blood. You will need the whole heart to be able to pump blood. Thus, the pumping property of the heart is an emergent or a supervenient property of the heart. Claiming that an individual heart cell can pump blood because the heart can would be an example of fallacy of division.
1. When we see complexification, that is: Interconnecting parts, weaving together, aggregating subunits, wrapping around, encompassing, interlinking, interlocking, twisting, interlacing, fusing, assembling related things, intricately combining things, where the system is greater than the sum of their parts. then it is logical to attribute such actions to intelligently acting mind with foresight and foreknowledge, and distant goals.
2. Making systems with the hallmark of complexity depends on the careful elaboration and design in detail of many elementary parts and interconnecting them in a meaningful way conferring a specific purpose or function. Not rarely, small changes in one part of the system can cause sudden and unexpected outputs in other parts of the system, system-wide reorganization, or breaking down of the higher function.
3. Random accidents are not the best case-adequate explanation for the origin of emerging properties of a complex system. intelligent design is.
Question: How could/would natural selection pressures operate, if there was no intent for these molecules to become part of living cells in a distant future?
Answer: Indeed, we find ourselves grappling with a profound question. As we delve into the realm of heterocycles and molecular structures, we must tread carefully, recognizing the limitations of metaphorical language. Nature, devoid of intent or decision-making capabilities, does not possess the faculties of choice or selection. The conundrum lies in the presence of specific heterocycles within the realms of biological systems—a puzzle that defies the boundaries of chemistry, physical necessity, and even the mechanisms of evolution. In the primordial days of our planet, a diverse array of conditions fostered the occurrence of chemical reactions. The Earth, with its mélange of elements and energy sources like lightning, volcanic activity, and the touch of UV radiation, served as a fertile crucible. Within this crucible, an assortment of chemical reactions unfolded, yielding a vast repertoire of molecules, including enigmatic heterocycles. However, we must pause and reflect upon the essence of natural selection, the supposed driving force that shapes and molds life as we know it. Within the context of early Earth's chemistry, there was no grand arbiter of selection, no force guiding the preservation of one molecule over another. The absence of higher-order systems poised for preservation or propagation leaves us pondering the fate of these molecular entities. Without a complex system to be favored and perpetuated, without the advantages of survival and competition, the notion of selection becomes a hollow echo. Thus, we confront the enigma of the molecular realm, particularly when it comes to the selection of molecules destined to become the bearers of information—such as DNA and RNA. Can we attribute this selection solely to the capricious whims of natural processes and chance? The intricate design and bewildering complexity that pervade these molecules suggest otherwise—they beckon us to consider the involvement of an intelligent creator. Nucleobases, those wondrous components that harbor the potential to store and transmit genetic information, engage in the delicate dance of base-pairing interactions, and facilitate the emergence of self-replicating systems, are bestowed with remarkable properties. These properties, far from emerging haphazardly, appear purposefully woven into the very fabric of their being—a tapestry of intentionality. As the quest for unguided mechanisms to explain the selection of life-permitting molecules continues, we find ourselves at an impasse—an impasse that beckons us to contemplate the alternative perspective of intelligent design. Within this perspective lies an acknowledgment of the profound complexity and deliberate arrangement that pervades the molecular realm, including the enigmatic nucleobases. Such intricate building blocks of life are best understood as the result of intentional design by a higher intelligence—an intelligence that transcends the bounds of the natural world. In our pursuit of understanding, we navigate the realms of science and philosophy, seeking glimpses of truth amidst the enigmatic foundation of existence. The intricate dance of molecules and their purposeful arrangement hints at a profound narrative—a narrative that invites us to explore the realms beyond naturalistic explanations and embrace the possibility of a guiding hand—an intelligence that shapes and breathes life into the very essence of our being.

Source: Pross A. (2021) Toward solving the mystery of the origin of life. Life, 11(9). https://www.mdpi.com/2075-1729/11/9/900
Calculations of life beginning through unguided, natural, random events.
http://reasonandscience.heavenforum.org/t2508-calculations-of-life-beginning-through-unguided-natural-random-events
1. https://webcache.googleusercontent.com/search?q=cache:Y7wv8TsUEQkJ:https://osf.io/7ke83/download/%3Fversion%3D4%26displayName%3DOrigin%2520of%2520Life%2520%2520%2520Stout%2520%2520Matzko%2520%25202018%2520%2520OSF-2019-05-05T05%253A50%253A03.408Z.pdf+&cd=1&hl=en&ct=clnk&gl=br
https://reasonandscience.catsboard.com/t2515-abiogenesis-the-possible-mechanisms-to-explain-the-origin-of-life
Either life just coalesced from atomic building blocks through a random fluke collision of disorderly pieces, emerging by “dumb, blind” mechanical processes, a fortuitous accident, spontaneously through self-organization by unguided, non-designed, unintended stochastic coincidence, natural events that turned into self-organization in an orderly manner without external direction, chemical non-biological, purely physico-dynamic kinetic processes and reactions influenced by environmental parameters, or through the direct intervention, direction-giving creative force, and design activity of an intelligent cognitive agency, a powerful conscious creator with intentions, inventive power, will, foreseeing goals and foresight, able to instantiate and create successful solutions in a planned manner.
Rational Wiki: Often brought up in the origins debate is how evolution does not explain the origin of life. Let's get something abundantly clear: abiogenesis and evolution are two completely different things. The theory of evolution says absolutely nothing about the origin of life. It merely describes the processes that take place once life has started.
https://rationalwiki.org/wiki/Abiogenesis
Koonin, E. V. (2012): The emergence of the first replicator system, which represented the “Darwinian breakthrough,” was inevitably preceded by a succession of complex, difficult steps for which biological evolutionary mechanisms were not accessible. The Logic of Chance: The Nature and Origin of Biological Evolution. Amazon.
ADDY PROSS (2012): Darwinian theory is a biological theory and therefore deals with biological systems, whereas the origin of life problem is a chemical problem, and chemical problems are best solved with chemical (and physical) theories. Attempting to explain chemical phenomena with biological concepts is methodologically problematic.
What is Life?: How Chemistry Becomes Biology
Paul Davies (2021): I think in all honesty a lot of people even confuse it the people who aren't familiar with the area that oh I presume Darwinian evolution sort of accounts for the origin of life but of course, you don't get an evolutionary process until you've got a self-replicating molecule. ( Darwin ) gave us a theory of evolution about how life has evolved but he uh didn't want to tangle with how you go from non-life to life and for me, that's a much bigger step. Why Darwinian evolution does NOT explain the origin of life
Fry, Iris. (2010): The Role of Natural Selection in the Origin of Life. Origins of Life and Evolution of Biospheres Link.
Iris Fry analyzes various theories on the origin of life, including RNA-first, metabolism-first, and others. She concludes that while none of these paradigms have decisive experimental support, gene-first theories show potential. As of her writing, no functioning system of genetic replication had been achieved without the addition of an external protein enzyme.
Alan W. Schwartz (2007): A problem that is familiar to organic chemists is the production of unwanted byproducts in synthetic reactions. For prebiotic chemistry, where the goal is often the simulation of conditions on the prebiotic Earth and the modeling of a spontaneous reaction, it is not surprising – but nevertheless frustrating – that the unwanted products may consume most of the starting material and lead to nothing more than an intractable mixture, or -gunk.. The most well-known examples of the phenomenon can be summarized quickly: Although the Miller –Urey reaction produces an impressive set of amino acids and other biologically significant compounds, a large fraction of the starting material goes into a brown, tar-like residue that remains uncharacterized; i.e., gunk. While 15% of the carbon can be traced to specific organic molecules, the rest seems to be largely intractable Even if we focus only on the soluble products, we still have to deal with an extremely complex mixture of compounds. The carbonaceous chondrites, which represent an alternative source of starting material for prebiotic chemistry on Earth, and must have added enormous quantities of organic material to the Earth at the end of the Late Heavy Bombardment (LHB), do not offer a solution to the problem just referred to. The organic material present in carbonaceous meteorites is a mixture of such complexity that much ingenuity has gone into the design of suitable extraction methods, to isolate the most important classes of soluble (or solubilized) components for analysis. Whatever the exact nature of an RNA precursor which may have become the first selfreplicating molecule, how could the chemical homogeneity which seems necessary to permit this kind of mechanism to even come into existence have been achieved? What mechanism would have selected for the incorporation of only threose, or ribose, or any particular building block, into short oligomers which might later have undergone chemically selective oligomerization? Virtually all model prebiotic syntheses produce mixtures. 2
A.G.Cairns-Smith (1985): It is one of the most singular features of the unity of biochemistry that this mere convention is universal. Where did such agreement come from? You see non-biological processes do not as a rule show any bias one way or the other, and it has proved particularly difficult to see any realistic way in which any of the constituents of a 'prebiotic soup' would have had predominantly 'left-handed' or right-handed' molecules. It is thus particularly difficult to see this feature as having been imposed by initial conditions. 3
A.G.Cairns-Smith (1985): genetic takeover, page 70: Suppose that by chance some particular coacervate droplet in a primordial ocean happened to have a set of catalysts, etc. that could convert carbon dioxide into D-glucose. Would this have been a major step forward towards life? Probably not. Sooner or later the droplet would have sunk to the bottom of the ocean and never have been heard of again. It would not have mattered how ingenious or life-like some early system was; if it lacked the ability to pass on to offspring the secret of its success then it might as well never have existed. So I do not see life as emerging as a matter of course from the general evolution of the cosmos, via chemical evolution, in one grand gradual process of complexification. Instead, following Muller (1929) and others, I would take a genetic View and see the origin of life as hinging on a rather precise technical puzzle. What would have been the easiest way that hereditary machinery could have formed on primitive Earth? 4
PROFESSOR DR KLAUS DOSE: The Origin of Life: More Questions Than Answers
Evolution, as used here, refers to any development or change influenced by the environment. The term "chemical evolution" specifically pertains to changes in chemical substances, indicating that fundamental transformations occur within molecules. Often, it is employed as a synonym for the "abiotic" or "prebiotic formation" of organic molecules in cosmic systems, particularly in the context of the early Earth. It is assumed that these organic molecules were produced from the constituents present in the primitive atmosphere, hydrosphere, and, to some extent, the lithosphere. On the other hand, "molecular evolution" encompasses a broader scope than chemical evolution. It encompasses self-assembly into more complex structures, such as membranes, protocells, cell-like systems, and protocellular organelles. Additionally, it involves the subsequent evolution of proto-cells or protobionts, leading to the emergence of the first modern cells, also known as "Urzellen" or progenotes. The transition from protobionts to progenotes has been referred to as proto-Darwinian evolution. In contrast, "Darwinian evolution" pertains to the evolution from progenotes to the vast array of contemporary cells and organisms.
https://sci-hub.ee/10.1179/isr.1988.13.4.348
Comment: According to the description provided, "chemical evolution" primarily focuses on the changes in chemical substances and the fundamental transformations that occur within molecules. It is often used as a term synonymous with the "abiotic" or "prebiotic formation" of organic molecules in cosmic systems, particularly in the context of the early Earth. The key aspect of chemical evolution is the formation of organic molecules from inorganic precursors found in the primitive atmosphere, hydrosphere, and lithosphere. Notably, the concept of "chemical evolution" does not explicitly include natural selection, which is a fundamental mechanism of Darwinian evolution. Natural selection is a process that acts on existing life forms, favoring certain traits or characteristics that provide a reproductive advantage in a given environment. Over time, this leads to the accumulation of beneficial traits within a population, leading to the adaptation and diversification of organisms. While chemical evolution sets the stage by forming the necessary building blocks of life, such as amino acids and nucleotides, it does not inherently involve the process of natural selection. Instead, it provides the foundation upon which subsequent evolutionary processes, like "molecular evolution" and "Darwinian evolution," can take place. "Molecular evolution," as mentioned in the initial description, goes beyond chemical evolution by encompassing the self-assembly of complex structures and the subsequent evolution of proto-cells to modern cells. This process involves changes in the molecular and structural organization of early life forms, leading to the emergence of more advanced cellular systems. On the other hand, "Darwinian evolution" encompasses the evolution of progenotes (early simple cells) to the vast array of contemporary cells and organisms we observe today. Natural selection is a central driving force in this form of evolution, shaping the diversity of life through the differential survival and reproduction of individuals with advantageous traits.
Self-organization refers to the supposed and hypothesized spontaneous emergence of order and complexity without external direction or control and without the need for external instructions. While self-organization is a hypothesized concept, it has never been directly observed or reproduced in a laboratory setting.
Gennady Shkliarevsky: THE UNIVERSAL EVOLUTION AND THE ORIGIN OF LIFE 2021
Most current OOL perspectives invoke chance or coincidence in their explanations of the origin of life. Sean Carroll, a well-known evolutionary biologist, refers to the emergence of life as “the mother of all accidents” and “the accident of all mothers.” There are also additional problems that plague the dominant scenario on the origin of prokaryotic cells. Most, if not all of them, invoke chance or coincidence to explain the origin of major cell components. Not only that, but they also invoke chance or coincidence to explain why these components came together to form a cell. As has been explained earlier, invoking chance even once is highly problematic; invoking it twice to explain the same phenomenon makes an explanation very questionable. As has been repeatedly pointed out, the liberal use of chance and coincidence as an explanatory mode is also a source of concern in theories about the origin and evolution of early life. In order to explain the emergence of radical novelty, many current OOL perspectives invoke chance or coincidence.
https://arxiv.org/ftp/arxiv/papers/2104/2104.08076.pdf
Alexei A. Sharov: Coenzyme world model of the origin of life 2016 Mar 9
The probability of transferring the full set of coding molecules to descendants by pure chance may be problematic especially if droplets carry too many kinds of coding molecules and some of them are present in a small number of copies. This combinatorial problem can be partially meliorated by the “stochastic corrector” mechanism, which is a preferential propagation of systems with a full set of coding molecules (Szathmáry, 1999). Systems with an incomplete set of coding molecules are more likely to fail in surviving and reproduction because some of their functions appear missing. This kind of stochastic correction is a primordial version of the purifying selection; and like the purifying selection, it reduces the overall reproduction rate of the population.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4875852/
Horst Rauchfuss Chemical Evolution and the Origin of Life
An important, but as yet unsolved, problem is provided by the chirality of the amino acids. The main question is whether the enantiomeric selection of the amino acids took place before or after the evolution of life.
Others see the genetic code as being purely fortuitous, a system that was “frozen” at some time in history.
William Dembski (2002): The problem is that nature has too many options and without design couldn’t sort through all those options. The problem is that natural mechanisms are too unspecific to determine any particular outcome. Natural processes could theoretically form a protein, but also compatible with the formation of a plethora of other molecular assemblages, most of which have no biological significance. Nature allows them full freedom of arrangement. Yet it’s precisely that freedom that makes nature unable to account for specified outcomes of small probability. Nature, in this case, rather than being intent on doing only one thing, is open to doing any number of things. Yet when one of those things is a highly improbable specified event, design becomes the more compelling, better inference. Occam's razor also boils down to an argument from ignorance: in the absence of better information, you use a heuristic to accept one hypothesis over the other. 5
There's nothing about inert chemicals and physical forces that say we want to get life at the end of the abiogenesis process. Molecules do not have the "drive", they do not "want" to find ways to harness the energy and become more efficient
One cannot explain the origin of evolution, through evolution. It is widespread and very common to see the attempt to smuggle the Darwinian dynamic of replication with a heritable variation into the origin of life. Biological evolution by natural selection does and cannot explain the origin of life. Natural selection only acts on the random variation of alleles based on DNA replication, but the origin of genes, and replication is among the origin of the entire self-replicating cell, what the origin of life research has to explain.
1. When we see complexification, that is: Interconnecting parts, weaving together, aggregating subunits, wrapping around, encompassing, interlinking, interlocking, twisting, interlacing, fusing, assembling related things, intricately combining things, where the system is greater than the sum of their parts. then it is logical to attribute such actions to an intelligently acting mind with foresight and foreknowledge, and distant goals.
2. Making systems with the hallmark of complexity depends on the careful elaboration and design in detail of many elementary parts and interconnecting them in a meaningful way conferring a specific purpose or function. Not rarely, small changes in one part of the system can cause sudden and unexpected outputs in other parts of the system, system-wide reorganization, or breaking down of the higher function.
3. Why WOULD a molecule self-replicate apart from duplication performed by the DNA replication machinery? There was no prebiotic natural selection with the goal to survive and reproduction. Why should be assumed that molecules - that are randomly assembled by chemical/physical forces/happenstance and just happened to stay in place - have the drive or goal to self-replicate, to become the complex macromolecular building blocks of living cells? Or to have any goal at all? Why would/should molecules strive toward increasing complexity without any possible "psychological" basis for doing so? Random accidents are not the best case-adequate explanation for the origin of emerging properties of a complex system. Intelligent design is.
Exploring the Deep Mystery of Life's Origins Aug 8, 2022
Nick Lane: Our kind of cell arose once in four billion years of evolution. And it seems to have been something of a bit of a freak accident.
https://www.youtube.com/watch?v=ATubwpnVLAY
Wilhelm T. S. Huck Robustness, Entrainment, and Hybridization in Dissipative Molecular Networks, and the Origin of Life May 30, 2019
Life emerged spontaneously from the selfassembly, or spontaneous organization, of the organic products of reactions, occurring in complex mixtures of molecules formed abiotically from simple precursors and sequences of reactions.
https://robobees.seas.harvard.edu/files/gmwgroup/files/1320.pdf
Phillip E. Johnson, DARWIN ON TRIAL: Darwin persuades us that the seemingly purposeful construction of living things can very often, and perhaps always, be attributed to the operation of natural selection.
If you have things that are reproducing their kind;
if there are sometimes random variations, nevertheless, in the offspring;
if such variations can be inherited;
if some such variations can sometimes confer an advantage on their owners;
if there is competition between the reproducing entities;-
if there is an overproduction so that not all will be able to produce offspring themselves-
then these entities will get better at reproducing their kind. What is needed for natural selection are things that conform to those 'ifs'. Self-replicating cells are prerequisites for evolution. None of this was available prebiotically to explain the origin of the first life form. 1
MARIO VANEECHOUTTE The scientific origin of life 2000
We hypothesize that the origin of life, that is, the origin of the first cell, cannot be explained by natural selection among self-replicating molecules, as is done by the RNA-world hypothesis.
The hypothesis espoused here states that it is virtually impossible that the highly complicated system cell developed gradually around simple self-replicating molecules (RNA-hypercycles or autocatalytic peptide networks) by means of natural selection; as is proposed by, for example, the RNA-world hypothesis. Despite searching quadrillions of molecules, it is clear that a spontaneous RNAreplicator is unlikely to occur. Reports of nucleotide and peptide self-replication still depend upon human intervention (for instance, by changing the environmental conditions between two rounds of replication or by denaturing the double strands). The problem of denaturing the double-nucleotide strand in a nonenzymatic manner has been overlooked and has contributed to a failure to establish molecular self-replication. The first cell, life, was born and natural selection (selection among variations on the theme of autonomous duplication) commenced.
The hypothesis suggested here states that no such autonomous duplication existed before the first cell and, thus, natural selection started only with the first cell.
https://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.645.3011&rep=rep1&type=pdf
Jonathan Sarfati Natural selection cannot explain the origin of life 31 March 2021
Natural selection requires pre-existing life.
https://creation.com/ns-origin-of-life
The first life could emerge by chance in a depression or tide pool, as one of countless reaction systems.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4810237/#:~:text=The%20bottom%2Dup%20approach%20is,%2C%20genetic%20code%2C%20and%20proteins.
Evolution by mutations and natural selection do not explain the origin of life, since evolution depends on the Darwinian dynamic of replication with a heritable variation. The only alternative to design are random unguided lucky events.
Or through the direct intervention and creative force of an intelligent agency, a powerful creator.
In an attempt to explain the origin of life, scientists propose a two-stage process of natural chemical evolution: formation of organic molecules, which combine to make larger biomolecules; self-organization of these molecules into a living organism. The origin of life can not be explained through biological nor chemical evolution. Adaptation, mutation, and natural selection depend on DNA replication. Heredity is guaranteed by faithful DNA replication whereas evolution depends upon errors accompanying DNA replication. Neither can it be explained through physical laws. Life depends on codes and instructional complex information. This information can only be generated by when the arrangement of the code is free and unconstrained, and any of the four bases of the genetic code can be placed in any of the positions in the sequence to generate the information. The only alternative, if the action of a creative agency is excluded, would be spontaneous self-assembly by orderly aggregation of prebiotic elements and building blocks in a sequentially correct manner without external direction.
Origin of life: A prebiotic route to DNA
The first polymeric molecules capable of storing information and reproducing themselves were randomly assembled from organic precursors that were available on the early Earth.
https://www.sciencedaily.com/releases/2019/06/190618103721.htm
Albert L. Lehninger described how inanimate matter crossed the great divide to become alive. “We now come to the critical moment in evolution,” he wrote in 1970, “in which the first semblance of ‘life’ appeared, through the chance association of a number of abiotically formed macromolecular components.”
Natural selection is not a possible mechanism to explain the origin of life, since evolution depends on DNA replication
Life's Origins: What Was the Origin Of Complex Organic Molecules? We are going to make an assumption: the chemical components found in all living prokaryotic and eukaryotic cells was present in the ancestral organism from which all life derived. In other words: the spontaneous interaction of the molecules present in the early earth's atmosphere formed more complex organic molecules, such as amino acids, sugars, fatty acids, and nitrogen bases - the building blocks of life as we know it today.
https://www2.gwu.edu/~darwin/BiSc151/Origin/origin.html
Question : Was there natural selection prior life starting?
Reply: No, natural selection as a mechanism of evolution cannot exist prior to the existence of life. Natural selection is a process by which certain traits that are better suited to an organism's environment become more prevalent in a population over time. This process depends on the existence of organisms that can reproduce, pass on traits to their offspring, and have variations in traits that can be selected for or against based on their survival and reproductive success.
In order for natural selection to occur, there must first be life, and this is where the question of the origin of life becomes relevant. While natural selection can explain the evolution of species once life has arisen, it cannot explain the initial emergence of life from non-living matter.
Stephen C. Meyer, The return of the God hypothesis
The process of natural selection presupposes the differential reproduction of already living organisms and thus a preexisting mechanism of self-replication. Yet self-replication in all extant cells depends upon functional (and therefore sequence-specific, information-rich) proteins and nucleic acids. And the origin of such information-rich molecules is precisely what Oparin needed to explain. Thus, many rejected his postulation of prebiotic natural selection as question-begging. As the evolutionary biologist Theodosius Dobzhansky insisted, “Pre-biological natural selection is a contradiction in terms.” Or as Christian de Duve explained, theories of prebiotic natural selection “need information which implies they have to presuppose what is to be explained in the first place.”
The Origin of the First Hereditary Replicators
This process is still an unsolved problem. By itself, this transition is not an evolutionary one because, without hereditary replicators, no Darwinian evolution is possible.
https://www.pnas.org/content/112/33/10104
Alemi Mario: The Amazing Journey of Reason from DNA to Artificial Intelligence 2020
Darwin probably didn’t propose a theory for the origin of life simply because applying Darwin’s mechanism of natural selection to the emergence of life, as done by Dawkins (1976), is like comparing apples with pears (Johnson 2010). What’s more, the idea that a self-replicating molecule with an information content casually appeared in a primordial soup, as imagined by Dawkins (1976) (“At some point a particularly remarkable molecule was formed by accident. We will call it the Replicator.”) appears to be statistically groundless (Yockey 1977).
https://link.springer.com/book/10.1007%2F978-3-030-25962-4
Is life a gamble? Scientist models universe to find out April 21, 2020
Scientists suspect that the complex life that slithers and crawls through every nook and cranny on Earth emerged from a random shuffling of non-living matter that ultimately spit out the building blocks of life.
https://www.livescience.com/origin-of-life-rna-universe-model.html
A. G. CAIRNS-SMITH Seven clues to the origin of life, page 36:
And if you ask me how the next stage happened, how the smallish 'molecules of life' came together to make the first reproducing evolving being, I will reply: 'With time, and more time, and the resource of oceans.' I will sweep my arms grandly about. 'Because you see. in the absence of oxygen the oceans would have accumulated "the molecules of life". The oceans would have been vast bowls of nutritious soup. Chance could do the rest.
The role of natural selection in the origin of life
Unlike living systems that are products of and participants in evolution, these prebiotic chemical structures were not products of evolution. Not being yet intricately organized, they could have emerged as a result of ordinary physical and chemical processes.
https://www.ncbi.nlm.nih.gov/pubmed/20407927
Alternative Pathways of Carbon Dioxide Fixation: Insights into the Early Evolution of Life? July 6, 2011
The fixation of inorganic carbon into organic material (autotrophy) is a prerequisite for life and sets the starting point of biological evolution.
https://sci-hub.ren/https://www.annualreviews.org/doi/10.1146/annurev-micro-090110-102801
Jack W. Szostak Functional proteins from a random-sequence library
Functional primordial proteins presumably originated from random sequences
https://molbio.mgh.harvard.edu/szostakweb/publications/Szostak_pdfs/Keefe_Szostak_Nature_01.pdf?fbclid=IwAR0giOg_aZfFRKQALk7CB22nVIx32ShiN0Vp78cwtAYwmwQ_0RJicfxpR1M
LIFE The Science of Biology, TENTH EDITION, page 3
When we consider how life might have arisen from nonliving matter, we must take into account the properties of the young Earth’s atmosphere, oceans, and climate, all of which were very different than they are today. Biologists postulate that complex biological molecules first arose through the random physical association of chemicals in that environment.
Neither Evolution nor physical necessity are a driving force prior dna replication. The only two alternatives are either a) creation by an intelligent agency, or b) Random, unguided, undirected natural events by a lucky "accident".
Koonin, the logic of chance, page 246
Evolution by natural selection and drift can begin only after replication with sufficient fidelity is established. Even at that stage, the evolution of translation remains highly problematic. The emergence of the first replicator system, which represented the “Darwinian breakthrough,” was inevitably preceded by a succession of complex, difficult steps for which biological evolutionary mechanisms were not accessible . The synthesis of nucleotides and (at least) moderate-sized polynucleotides could not have evolved biologically and must have emerged abiogenically—that is, effectively by chance abetted by chemical selection, such as the preferential survival of stable RNA species. Translation is thought to have evolved later via an ad hoc selective process. Did you read this ???!! An ad-hoc process ??
Without code there can be no self-replication. Without self-replication, you can’t have reproduction. Without reproduction, you can’t have evolution or natural selection.
Heredity is guaranteed by faithful DNA replication whereas evolution depends upon errors accompanying DNA replication. ( Furusawa, 1998 ) We hypothesize that the origin of life, that is, the origin of the first cell, cannot be explained by natural selection among self-replicating molecules, as is done by the RNA-world hypothesis. ( Vaneechoutte M )
Chance and necessity do not explain the origin of life
https://www.academia.edu/1204161/Trevors_J.T._Abel_D.L._2004_Chance_and_necessity_do_not_explain_the_origin_of_life_Cell_Biology_International_28_729-739
Selection pressure cannot select nucleotides at the digital programming level where primary structures form. Genomes predetermine the phenotypes which natural selection only secondarily favors. Contentions that offer nothing more than long periods of time offer no mechanism of explanation for the derivation of genetic programming. No new informationis provided by such tautologies. The argument simply says it happened. As such, it is nothing more than blind belief. Science must provide rational theoretical mechanism, empirical support, prediction fulfillment, or some combination of these three. If none of these three are available, science should reconsider that molecular evolution of genetic cybernetics is a proven fact and press forward with new research approaches which are not obvious at this time. 5
I would like to plead with you, simply, please realize you cannot use the words `natural selection' loosely. Prebiological natural selection is a contradiction of terms."
(Dobzhansky, T.G., Discussion of "Synthesis of Nucleosides and Polynucleotides with Metaphoric Esters,", Oct. 27-30, 1963, Academic Press: New York NY, 1965, pp.309-310).
B.Alberts, Molecular Biology of the Cell, 5th edition, page 406
Self-Replicating Molecules Undergo Natural Selection
The three-dimensional folded structure of a polynucleotide affects its stability, its actions on other molecules, and its ability to replicate. Therefore, certain polynucleotides will be especially successful in any self-replicating mixture. Because errors inevitably occur in any copying process, new variant sequences of these polynucleotides will be generated over time.

Stephen Meyer, Darwin's doubt, page 6:
Natural selection assumes the existence of living organisms with a capacity to reproduce. Yet self-replication in all extant cells depends upon information-rich proteins and nucleic acids (DNA and RNA), and the origin of such information-rich molecules is precisely what origin-of-life research needs to explain. That’s why Theodosius Dobzhansky, one of the founders of the modern neo-Darwinian synthesis, can state flatly, “Pre-biological natural selection is a contradiction in terms.”5 Or, as Nobel Prize–winning molecular biologist and origin-of-life researcher Christian de Duve explains, theories of prebiotic natural selection fail because they “need information which implies they have to presuppose what is to be explained in the first place.
That means, evolution was not a driving force and acting for the emergence and origin of the first living organisms. The only remaining possible mechanisms are chemical reactions acting upon unregulated, aleatory events ( luck, chance), or physical necessity. ( where chemical reactions are forced into taking a certain course of action. )
Morowitz: THE ORIGIN AND NATURE OF LIFE ON EARTH page 18
The Darwinian framework for selection requires support from other error-correcting mechanisms that operate in simpler contexts, to arrive at a mechanism sufficient to explain the emergence, overall organization, and long-term persistence of life from non-living precursors.
In a relatively short time, the ocean became a broth of these molecules, and given enough time, the right combination of molecules came together by pure chance to form a replicating entity of some kind that evolved into modern life.
https://www.americanscientist.org/article/the-origin-of-life
Physical necessity & Physical laws
Stephen C.Meyer, The return of the God hypothesis, page 216:
Rather than having a genetic molecule capable of unlimited novelty, with all the unpredictable and aperiodic sequences that characterize informative texts, we would have a highly repetitive text awash in redundant sequences—much as happens in crystals. Indeed, in a crystal the forces of mutual chemical attraction do completely explain the sequential ordering of the constituent parts. Consequently, crystals cannot convey novel information. Bonding affinities, to the extent they exist, cannot be used to explain the origin of information. Self-organizing chemical affinities generate highly repetitive “order,” but not information; they create mantras, not messages
The nucleotide sequence of DNA and RNA have an instructional function to make proteins and is NOT random but complex and specified, and not due to physical necessity or physical laws. And this is what events in a prebiotic land would need to produce: a minimal set of proteins .... and this kind of specification does not arise through chemical reactions ...... the result of a chemical reaction is not random. But the events dealing with an eventual chemical reaction would have been if there was not a mind guiding the events.
Consider, for example, what would happen if the individual nucleotide bases (A, C, G, T) in the DNA molecule did interact by chemical necessity (along the information-bearing axis of DNA). Suppose that every time adenine (A) occurred in a growing genetic sequence, it attracted cytosine (C) to it,26 which attracted guanine (G), which attracted thymine (T), which attracted adenine (A), and so on. If this were the case, the longitudinal axis of DNA would be peppered with repetitive sequences of ACGT. Rather than being a genetic molecule capable of virtually unlimited novelty and characterized by unpredictable and aperiodic sequences, DNA would contain sequences awash in repetition or redundancy—much like the arrangement of atoms in crystals.
Michael Polanyi: Life's Irreducible Structure,
Science mag, 1968
In Galileo's experiments on balls rolling down a slope, the angle of the slope was not derived from the laws of mechanics, but was chosen by Galileo. And as this choice of slopes was extraneous to the laws of mechanics, so is the shape and manufacture of test tubes extraneous to the laws of chemistry. The same thing holds for machinelike boundaries; their structure cannot be defined in terms of the laws which they harness. Nor can a vocabulary determine the content of a text, and so on. Therefore, if the structure of living things is a set of boundary conditions, this structure is extraneous to the laws of physics and chemistry which the organism is harnessing. Thus the morphology of living things transcends the laws of physics and chemistry.the codelike structure of DNA must be assumed to have come about by a sequence of chance variations established by natural selection. But this evolutionary aspect is irrelevant here; whatever may be the origin of a DNA configuration, it can function as a code only if its order is not due to the forces of potential energy. It must be as physically indeterminate as the sequence of words is on a printed page. As the arrangement of a printed page is extraneous to the chemistry of the printed page, so is the base sequence in a DNA molecule extraneous to the chemical forces at work in the DNA molecule. It is this physical indeterminacy of the sequence that produces the improbability of occurrence of any particular sequence and thereby enables it to have a meaning-a meaning that has a mathematically determinate information content equal to the numerical improbability of the arrangement.
A deterministic answer assumes that the laws of physics and chemistry have causally and sequentially determined the obligatory series of events leading from inanimate matter to life – that each step is causally linked to the previous one and to the next one by the laws of nature. In principle, in a strictly deterministic situation, the state of a system at any point in time determines the future behavior of the system – with no random influences. To invoke a guided determinism toward the formation of life would only make sense if the construction of life was demonstrably a preferential, highly probable natural pathway.
Luisi, The Emergence of Life; From Chemical Origins to Synthetic Biology, page 21
Just like computer codes, the genetic code is arbitrary. There is no law of physics that says “1” has to mean “on” and “0” has to mean “off.” There’s no law of physics that says 10000001 has to code for the letter “A.” Similarly, there is no law of physics that says three Guanine molecules in a row have to code for Glycine. In both cases, the communication system operates from a freely chosen, fixed set of rules.
In all communication systems it is possible to label the encoder, the message and the decoder and determine the rules of the code.
The rules of communication systems are defined in advance by conscious minds. There are no known exceptions to this. Therefore we have 100% inference that the Genetic Code was designed by a conscious mind.
Physical laws which result in physical constraints, where chemical reactions are forced into taking a certain course of action is an often cited possible mechanism for the origin of life.
We are moving from chemistry to biology. Henceforward, life, it goes without saying, is independent of its chemical substrate, and its evolution does not follow paths that are predictable solely based on the laws of physics.
M. Gargaud · H. Martin · P. López-García T. Montmerle · R. Pascal Young Sun, Early Earth and the Origins of Life, page 95
Laurent Boiteau Prebiotic Chemistry: From Simple Amphiphiles to Protocell Models, page 3:
Spontaneous self-assembly occurs when certain compounds associate through noncovalent hydrogen bonds, electrostatic forces, and nonpolar interactions that stabilize orderly arrangements of small and large molecules. The argument that chemical reactions in a primordial soup would not act upon pure chance, and that chemistry is not a matter of "random chance and coincidence, finds its refutation by the fact that the information stored in DNA is not constrained by chemistry. Yockey shows that the rules of any communication system are not derivable from the laws of physics. He continues: “there is nothing in the physicochemical world that remotely resembles reactions being determined by a sequence and codes between sequences.” In other words, nothing in nonliving physics or chemistry obeys symbolic instructions.
Ulrich E. Stegmann: The arbitrariness of the genetic code March 2004 5
Some of the processes expected to involve semantic information are certainly not chemically arbitrary and, therefore, chemical arbitrariness is not a necessary condition for a semantic relation.
The problem of information to explain the origin of life
Norbert Weiner - MIT Mathematician - Father of Cybernetics
"Information is information, not matter or energy. No materialism which does not admit this can survive at the present day."
It has to be explained:
- a library index and fully automated information classification, storage and retrieval program ( chromosomes, and the gene regulatory network )
- The origin of the complex, codified, specified, instructional information stored in the genome and epigenetic codes to make the first living organism
- The origin of the genetic Code
- How it got nearly optimal for allowing additional information within protein-coding sequences
- How it got more robust than 1 million alternative possible codes
- The origin of the over twentythree epigenetic codes
- The origin of the information transmission system, that is the origin of the genetic code itself, encoding, transmission, decoding and translation
- The origin of the genetic cipher/translation, from digital ( DNA / mRNA ) to analog ( Protein )
- The origin of the hardware, that is DNA, RNA, amino acids, and carbohydrates for fuel generation
- The origin of the replication/duplication of the DNA
- The origin of the signal recognition particle
- The origin of the tubulin Code for correct direction to the final destination of proteins
Information theory cannot normally be used to predict how chemicals will react because some chemicals react with each other readily, and others only react very slowly. Others do not react with each other at all. Thus, the likelihood of two chemicals joining together depends on both the quantity of the chemicals present and their chemical properties. Information theory can easily deal with the effects of quantity, but it has no way to deal with chemical properties.
Stuart Pullen, Intelligent Design or Evolution? Why the Origin of Life and the Evolution of Molecular Knowledge Imply Design, page 88
http://lifesorigin.com/prebiotic-evolution4.pdf
Stephen C. Meyer observed:
“There are neither bonds nor bonding affinities—differing in strength or otherwise—that can explain the origin of the base sequencing that constitutes the information in the DNA molecule”
(Signature in the Cell, 243).
As Paul Davies lamented,
“We are still left with the mystery of where biological information comes from.… If the normal laws of physics can’t inject information, and if we are ruling out miracles, then how can life be predetermined and inevitable rather than a freak accident? How is it possible to generate random complexity and specificity together in a lawlike manner? We always come back to that basic paradox”
(Fifth Miracle, 258).
A law of nature could not alone explain how life began, because no conceivable law would compel a legion of atoms to follow precisely a prescribed sequence of assemblage.
Paul Davies, The origin of Life, page 17
Werner Gitt summarized it this way:
“A necessary requirement for generating meaningful information is the ability to select from alternatives and this requires an intelligent, volitional entity.… Unguided, random processes cannot do this—not in any amount of time because this selection process demands continuous guidance by intelligent beings that have a purpose”
(Without Excuse, 50–51).
Let's assume that we begin with the sequence R-T-X, and will add two amino acids "B" and "A" to it. If amino acid "B" is the most reactive amino acid, the sequence would be R-T-X-B-A. However, if "A" is the most reactive amino acid, then the sequence would be R-T-X-A-B. In a random chemical reaction, the sequence of amino acids would be determined by the relative reactivity of the different amino acids. The polymer chain found in natural proteins and DNA has a very precise sequence that does not correlate with the individual components' reaction rates. Since all of the amino acids have relatively similar structures, they all have similar reaction rates; they will all react at about the same rate making the precise sequence by random chemical reactions unthinkably unlikely. This is the problem of Chemical Reactivity.
http://www.icr.org/article/evolution-hopes-you-dont-know-chemistry-problem-co/
The Genetic Code
http://hyperphysics.phy-astr.gsu.edu/hbase/Organic/gencode.html
DNA contains a true code. Being a true code means that the code is free and unconstrained; any of the four bases can be placed in any of the positions in the sequence of bases. Their sequence is not determined by the chemical bonding. There are hydrogen bonds between the base pairs and each base is bonded to the sugar phosphate backbone, but there are no bonds along the longitudional axis of DNA. The bases occur in the complementary base pairs A-T and G-C, but along the sequence on one side the bases can occur in any order, like the letters of a language used to compose words and sentences. Since nucleotides can be arranged freely into any informational sequence, physical necessity could not be a driving mechanism.
Abiogenesis is the process by which life arises naturally from non-living matter. Scientists speculate that life may have arisen as a result of random chemical processes happening to produce self-replicating molecules.
http://rationalwiki.org/wiki/Abiogenesis
Paul Davies conceded, “Unfortunately, before Darwinian evolution can start, a certain minimum level of complexity is required. But how was this initial complexity achieved? When pressed, most scientists wring their hands and mutter the incantation ‘Chance.’ So, did chance alone create the first self-replicating molecule?” (Fifth Miracle, 138).
If design or physical necessity is discarded, the only remaining possible mechanism for the origin of life is chance/luck.
Emergent properties
Claim: The origin of life could be an emergent property of basic chemical reactions
Reply: An emergent property is a property that a collection or complex system has, but which the individual members do not have. In biology, for example, heart is made of heart cells, heart cells on their own don't have the property of pumping blood. You will need the whole heart to be able to pump blood. Thus, the pumping property of the heart is an emergent or a supervenient property of the heart. Claiming that an individual heart cell can pump blood because the heart can would be an example of fallacy of division.
1. When we see complexification, that is: Interconnecting parts, weaving together, aggregating subunits, wrapping around, encompassing, interlinking, interlocking, twisting, interlacing, fusing, assembling related things, intricately combining things, where the system is greater than the sum of their parts. then it is logical to attribute such actions to intelligently acting mind with foresight and foreknowledge, and distant goals.
2. Making systems with the hallmark of complexity depends on the careful elaboration and design in detail of many elementary parts and interconnecting them in a meaningful way conferring a specific purpose or function. Not rarely, small changes in one part of the system can cause sudden and unexpected outputs in other parts of the system, system-wide reorganization, or breaking down of the higher function.
3. Random accidents are not the best case-adequate explanation for the origin of emerging properties of a complex system. intelligent design is.
Question: How could/would natural selection pressures operate, if there was no intent for these molecules to become part of living cells in a distant future?
Answer: Indeed, we find ourselves grappling with a profound question. As we delve into the realm of heterocycles and molecular structures, we must tread carefully, recognizing the limitations of metaphorical language. Nature, devoid of intent or decision-making capabilities, does not possess the faculties of choice or selection. The conundrum lies in the presence of specific heterocycles within the realms of biological systems—a puzzle that defies the boundaries of chemistry, physical necessity, and even the mechanisms of evolution. In the primordial days of our planet, a diverse array of conditions fostered the occurrence of chemical reactions. The Earth, with its mélange of elements and energy sources like lightning, volcanic activity, and the touch of UV radiation, served as a fertile crucible. Within this crucible, an assortment of chemical reactions unfolded, yielding a vast repertoire of molecules, including enigmatic heterocycles. However, we must pause and reflect upon the essence of natural selection, the supposed driving force that shapes and molds life as we know it. Within the context of early Earth's chemistry, there was no grand arbiter of selection, no force guiding the preservation of one molecule over another. The absence of higher-order systems poised for preservation or propagation leaves us pondering the fate of these molecular entities. Without a complex system to be favored and perpetuated, without the advantages of survival and competition, the notion of selection becomes a hollow echo. Thus, we confront the enigma of the molecular realm, particularly when it comes to the selection of molecules destined to become the bearers of information—such as DNA and RNA. Can we attribute this selection solely to the capricious whims of natural processes and chance? The intricate design and bewildering complexity that pervade these molecules suggest otherwise—they beckon us to consider the involvement of an intelligent creator. Nucleobases, those wondrous components that harbor the potential to store and transmit genetic information, engage in the delicate dance of base-pairing interactions, and facilitate the emergence of self-replicating systems, are bestowed with remarkable properties. These properties, far from emerging haphazardly, appear purposefully woven into the very fabric of their being—a tapestry of intentionality. As the quest for unguided mechanisms to explain the selection of life-permitting molecules continues, we find ourselves at an impasse—an impasse that beckons us to contemplate the alternative perspective of intelligent design. Within this perspective lies an acknowledgment of the profound complexity and deliberate arrangement that pervades the molecular realm, including the enigmatic nucleobases. Such intricate building blocks of life are best understood as the result of intentional design by a higher intelligence—an intelligence that transcends the bounds of the natural world. In our pursuit of understanding, we navigate the realms of science and philosophy, seeking glimpses of truth amidst the enigmatic foundation of existence. The intricate dance of molecules and their purposeful arrangement hints at a profound narrative—a narrative that invites us to explore the realms beyond naturalistic explanations and embrace the possibility of a guiding hand—an intelligence that shapes and breathes life into the very essence of our being.


Source: Deamer D.W. and Damer B. (2017) Can Life Begin on Enceladus? A Perspective from Hydrothermal Chemistry. Astrobiology, 17(9). https://www.liebertpub.com/doi/10.1089/ast.2016.1588How might life have originated on the early Earth? In 1859, Charles Darwin speculated that life might have begun in a “warm little pond” but most research since has focused on understanding how the basic chemicals of life might have formed. There was almost no investigation of how the first life forms appeared and then evolved into the complex biochemical systems we see today.
Abiogenesis is a historical event that cannot be reproduced experimentally. Therefore, the pathways leading to the origin of life remain unknown.
Source: Pross A. (2021) Toward solving the mystery of the origin of life. Life, 11(9). https://www.mdpi.com/2075-1729/11/9/900
Source: Wickramasinghe C. (2015) The search for our cosmic ancestry. World Scientific.The emergence of life from inanimate matter was a singularity, a unique event in history, which physics and chemistry cannot explain.
Source: Ruiz-Mirazo K., Briones C., and de la Escosura A. (2014) Prebiotic Systems Chemistry: New Perspectives for the Origins of Life. Chemical Reviews, 114(1). https://pubs.acs.org/doi/10.1021/cr2004844How life emerged from nonlife remains a fundamental unsolved problem.
Source: Trevors J.T. and Pollack G.H. (2012) Chance and necessity do not explain the origin of life. Cell Biology International, 36(7). https://onlinelibrary.wiley.com/doi/10.1042/CBI20110348There is no generally accepted description of how life on Earth originated. All existing theories of abiogenesis fall short of completeness.
Calculations of life beginning through unguided, natural, random events.
http://reasonandscience.heavenforum.org/t2508-calculations-of-life-beginning-through-unguided-natural-random-events
1. https://webcache.googleusercontent.com/search?q=cache:Y7wv8TsUEQkJ:https://osf.io/7ke83/download/%3Fversion%3D4%26displayName%3DOrigin%2520of%2520Life%2520%2520%2520Stout%2520%2520Matzko%2520%25202018%2520%2520OSF-2019-05-05T05%253A50%253A03.408Z.pdf+&cd=1&hl=en&ct=clnk&gl=br
Last edited by Otangelo on Fri 4 Oct 2024 - 17:55; edited 110 times in total
