Evolution of fish
http://reasonandscience.heavenforum.org/t2482-evolution-of-fish
“As we move back down our evolutionary ladder, jaw structure becomes more instead of less complex, and in fishes the jaws are very elaborate indeed” (Maisey, 2000, p. 61).
Here we enter a strange and poorly known area of phylospace, one in which both timing and phylogeny are poorly understood. Somewhere between 750 and 550 million years ago, a new line of chordates evolved. These were characterised by a relatively concentrated set of sensory neurons at the anterior end and a particularly pronounced notochord extending beyond the anterior and posterior ends of the torso. In other words, they had the beginnings of a head and a tail. In addition, probably as a later development, the myomeres, the charcateristic bands of muscle along the torso, were reorganized in dorsal and ventral halves, separated by a septum, and acquired a distinctive 'V'- or 'W'-shape. These folks were our ancestors among chordate-kind. 4
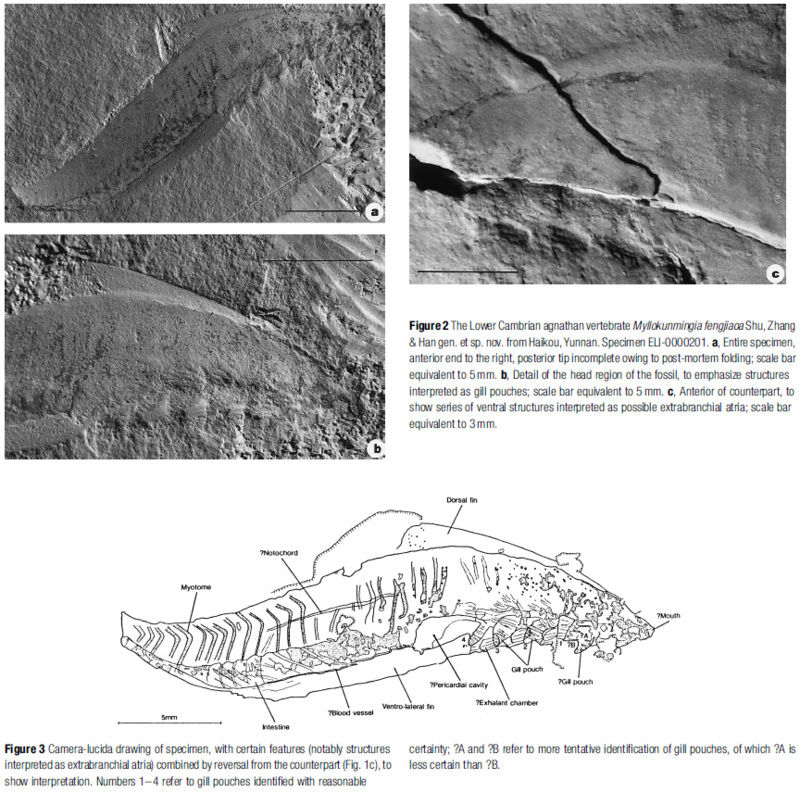
The last common ancestor of hagfish and you, with all of its descendants, is referred to as Craniata. Craniates are animals with well-developed heads and a skull of sorts. A very early craniate, Myllokunmingia from the Cambrian of China, is badly illustrated in the figure at right. A rather better illustration of a possibly related and similar form is shown here. Since there is no really good reason to suppose that Haikouella belonged to this clade, we treat as a sort of "craniatomorph," but not a craniate.
From the first craniates evolved a bewildering variety of creatures. At least they have bewildered systematists for over a century. The first step seems relatively clear. The most primitive craniates we know are the living Myxinoidea (hagfishes): eel-like, rather assymmetric creatures with a strong propensity to flood their immediate neighborhood with slime at the slightest provocation. All other known craniates are Vertebrates. ATW020417.
There are three subphylums in chordata
Urochordata (tunicates)
Cephalochordata (lancelets)
Vertebrata
The First Vertebrates, Jawless Fishes, the Agnathans
Vertebrates arose over 500 million years ago (MYA) but traces of their appearance only occur during the Ordovician period about 460 MYA. These primeval, small fish-like vertebrates are popularly known as the ostracoderms and during the 100 million years of their existence they were comprised of about 600 species. Ostracoderms are especially important in the history and evolution of vertebrates. As undoubted vertebrates they possessed a backbone, were bilaterally symmetrical and had a nervous system divided into brain and spinal cord (partly enclosed within the backbone). They are also characterized by the possession of no more than two pairs of limbs and muscular system consisting primarily of bilaterally paired masses and a well-developed coelom, which contained the organs. In appearance, ostracoderms were dorsoventrally flattened and, quite extraordinarily, they lacked jaws, a condition so important that the classification of modern vertebrates is recognized by two major groups, the
agnathans (without jaws) and the
gnathostomes (literally, jaw-mouths)
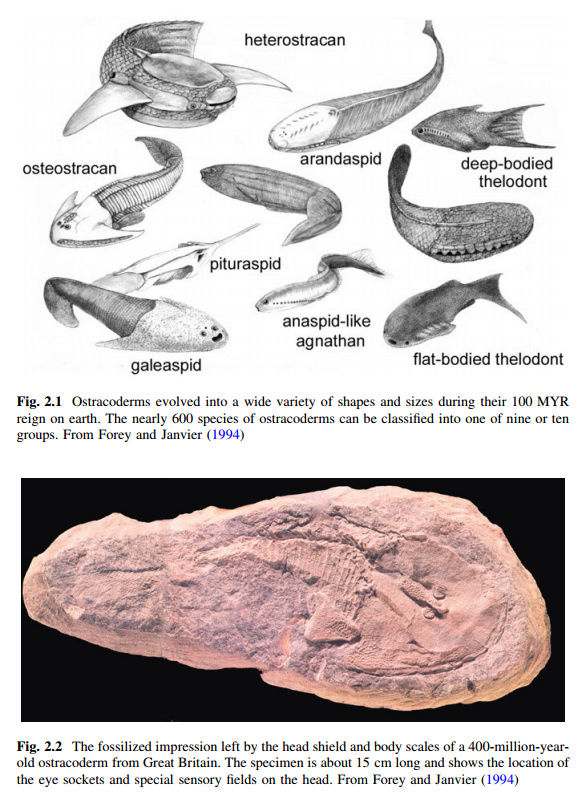
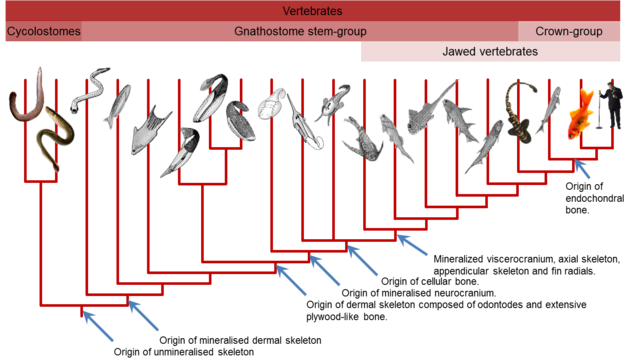
Ostracoderms are now regarded as an artificial designation that includes perhaps four distinct superclasses of jawless craniate fishes, the Pteraspidomorphi, Anaspida, Thelodonti, and Osteostrachomorphi. Despite an apparently inauspicious beginning, these simple animals would be responsible for two events of major importance in the history of the vertebrates—the evolution of a tough outer protective layer, the integument, and the evolution of bone. Remarkably, in the beginning the histories of these seemingly disparate structures were intrinsically linked. Ostracoderms possessed an external bony head shield or armor but its internal skeleton was probably not ossified to any great extent. Debate over the origin of the vertebrate skeleton has revolved around the question of which group was the first to exhibit evidence of skeletonization and mineralization. The vertebrate skeleton is comprised of at least two distinct skeletal systems: the dermoskeleton (arising from the dermis; widely perceived to encompass teeth, scales, fin spines, etc.) and the endoskeleton (the braincase, branchial skeleton, axial, and appendicular ). It has been argued that although elements of the skeleton can be interchangeably derived from either system, the two systems have remained distinct throughout vertebrate phylogeny (Donoghue and Sansom 2002), of which more will be said later (Fig. 2.2).
Ostracoderms possessed an external bony head shield or armor but its internal skeleton was probably not ossified to any great extent. Debate over the origin of the vertebrate skeleton has revolved around the question of which group was the first to exhibit evidence of skeletonization and mineralization. The vertebrate skeleton is comprised of at least two distinct skeletal systems:
the dermoskeleton (arising from the dermis; widely perceived to encompass teeth, scales, fin spines, etc.) and the
endoskeleton (the braincase, branchial skeleton, axial, and appendicular skeletons).
It has been argued that although elements of the skeleton can be interchangeably derived from either system, the two systems have remained distinct throughout vertebrate phylogeny (Donoghue and Sansom 2002) (Fig. 2.2).
The evolution of fish began about 530 million years ago during the Cambrian explosion. According to the theory of evolution, life began in the seas, and the first advanced vertebrate animals were fish. Again according to the theory, these fish began to move toward dry land and in some way, came to use feet instead of fins and lungs instead of gills! Many books on evolution never consider the how of this major claim, whose baselessness is glossed over in most scientific textbooks in some summary like ". . . and living things moved from the water to dry land."
“A history of fishes” 2
Fishes were the first vertebrates. Understanding theevolutionary history origin of fishes is therefore important not only for what it tells us about fish groups, but for what it tells us about evolution of the vertebrates and ultimately our own species. Innovations during fish evolution that were passed on to higher vertebrates include dermal and endochondral bone and their derivatives (vertebral centra, bony endoskeletons, brain cases, teeth), jaws, brains, appendages, and the internal organ systems that characterize all vertebrate groups today. During 500 million years of evolution, fishes colonized and dominated the seas and fresh waters and eventually emerged, at least for short periods, onto land . ( " Tetrapods evolved " . Really ? ) Major clades prospered and vanished, or were replaced by newer groups with presumably superior innovations. Extant (“living”) fishes therefore represent the most recent manifestations of adaptations and lineages that have their roots in the early Paleozoic. The more than 27,000 species of extant fishes constitute only a fraction of the diversity of fishes that has existed historically, as should be evident from the long lists of extinct forms given here (which in turn represent a select fraction of the diversity of former taxa). Many of the extinct forms are exotic in their appearance, whereas others are remarkably similar to living forms, at least in external morphology. A major challenge to ichthyology involves unraveling the evolutionary pathways of both modern and past fish taxa in the process of determining relationships among groups. Which of the many fossil groups represent ancestral types? Which were independent lineages that died out without representation in modern forms? What are the links between and among groups of the past and present? What do fossilized traits tell us about ancient environments? Where do similarities represent inheritance, convergence, or coincidence among extinct and living groups? And how have past adaptations influenced and perhaps constrained present morphologies and behaviors?
The focus is on fishes that lived during the Paleozoic and Mesozoic eras, and on modifications that occurred during the evolution of different, major extinct groups, leading to the dominant bony and cartilaginous fishes of today. We deal first with jawless fishes, then with ancestors of modern bony fishes because these occur earlier in the fossil record, and finally with the cartilaginous sharks, skates, rays, and chimaeras. This presentation focuses on extinct rather than extant fishes, recognizing that the distinctionb is artificial, that many lineages arose hundreds of millions of years ago and still have modern, living representatives, and that direct ancestors of some extant forms arose before other groups that have since gone extinct . We follow the basic organization of Nelson (2006) because of its synthetic and broad approach, recognizing that Nelson’s conclusions are one of many alternative interpretations of the literature.
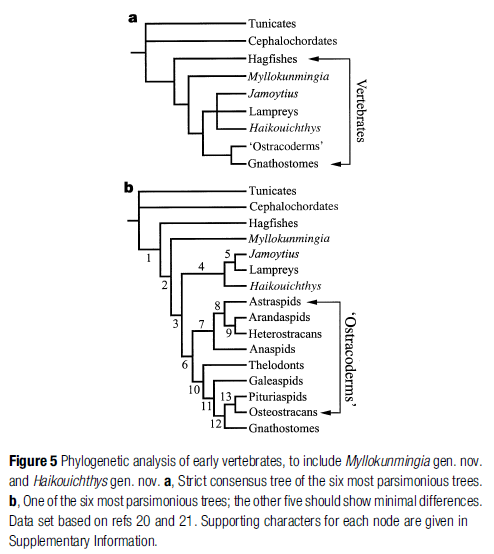
The very first fishlike vertebrates undoubtedly evolved from invertebrates, perhaps a cephalochordate ( how do they know this is in sequence they admit that : ). However, the first “fishes” left no fossil record and their form and relationships remain a mystery. By the time fishlike fossils appear in Early Cambrian deposits, roughly 530 million years before present (Fig. 11.1), complex tissue types had evolved, including filamentous gills, V-shaped myomeres, and a distinct dorsal fin. New discoveries are made almost annually, but the currently recognized oldest species, Myllokunmingia fengjiaoa, was found in the Chengjiang geological formation of Yunnan Province in southwestern China. Myllokunmingia was 3– 4 cm long and is thought to be allied with (is a sister group to) ancestors of modern lampreys, although agreement is far from universal (Xian-guang et al. 2002;
Shu et al. 2003).
Lower Cambrian vertebrates from south China
NATURE | VOL 402 | 4 NOVEMBER 1999 |
The first fossil chordates are found in deposits from the Cambrian period (545±490 million years ago), but their earliest record is exceptionally sporadic and is often controversial.
Chordate is an animal belonging to the phylum Chordata; they possess a notochord, a hollow dorsal nerve cord, pharyngeal slits, an endostyle, and a post-anal tail, for at least some period of their life cycle. Chordates are deuterostomes, as during the embryo development stage the anus forms before the mouth. They are also bilaterally symmetric coelomates. In the case of vertebrate chordates, the notochord is usually replaced by a vertebral column during development, and they may have body plans organized via segmentation.
Accordingly, it has been diffcult to construct a coherent phylogenetic synthesis for the basal chordates. Until now, the available soft-bodied remains have consisted almost entirely of cephalochordate-like animals from Burgess Shale-type faunas. Definite examples of agnathan fish do not occur until the Lower Ordovician (,475 Myr BP), with a more questionable record extending into the Cambrian. The discovery of two distinct types of agnathan from the Lower Cambrian Chengjiang fossil-Lagerstaette is, therefore, a very significant extension of their range. One form is lampreylike, whereas the other is closer to the more primitive hagfish. These finds imply that the first agnathans may have evolved in the earliest Cambrian, with the chordates arising from more primitive deuterostomes in Ediacaran times (latest Neoproterozoic,,555 Myr BP), if not earlier.
If modern cephalochordates such as the lancelets (Branchiostoma) are considered fishlike
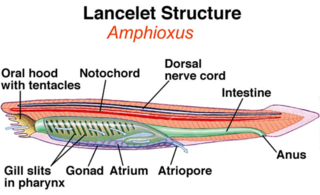
– if not exactly fishes –then the ancestry of fishes can be traced farther back to the cephalochordate-like yunnanozoans (Haikouella and Yunnanozoon) from the Lower Cambrian, or to the muchheralded Pikaia with its dorsal nerve cord and notochord, from the Middle Cambrian Burgess Shale of British Columbia. Clearly recognizable fish specimens, such as the arandaspid pteraspidomorph Sacabambaspis janvieri from Bolivia, appear later, dating to 470 mybp (Gagnier et al. 1986; Gagnier 1989). This and related jawless, finless forms inhabited shallow seas or estuarine habitats in tropical and subtropical regions of the Gondwanan and Laurasian supercontinents. Their innovations include true bone (probably evolved independently in several ancestral groups) and a muscular feeding pump. The former adaptation, which existed only as an external covering, would have provided protection from predators to which softer ancestors were more vulnerable, as well as serving as a metabolic reserve for calcium and phosphate and an insulator of electrosensory organs (Northcutt & Gans 1983; Carroll 1988). A muscular feeding pump would have been more efficient for moving food-bearing water through a filtration mechanism than was the ciliary feeding mechanism of protochordates. Another major advance over the cephalochordates that preceded them was that, although lacking jaws, the early fossilized fishes were craniates. They had a head region containing a brain with specialized sensory capsules and cranial nerves, all contained in a protective skeletal braincase (Maisey 1996).
Subphylum Craniata
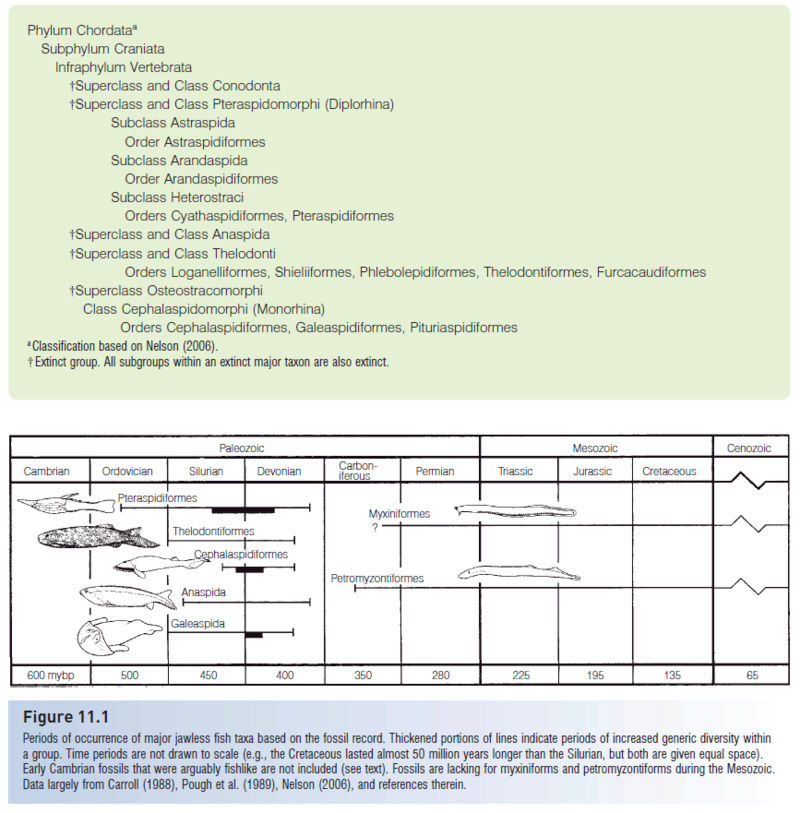
Vertebrate craniates possess, among other features, a dermal skeleton and neural crest, the latter describing regions of the developing nerve cord that are precursors to gill arches, pigment cells, connective tissue, and bone. Within the vertebrates are seven superclasses of fishes, five of which are extinct.
†Pteraspidomorphi
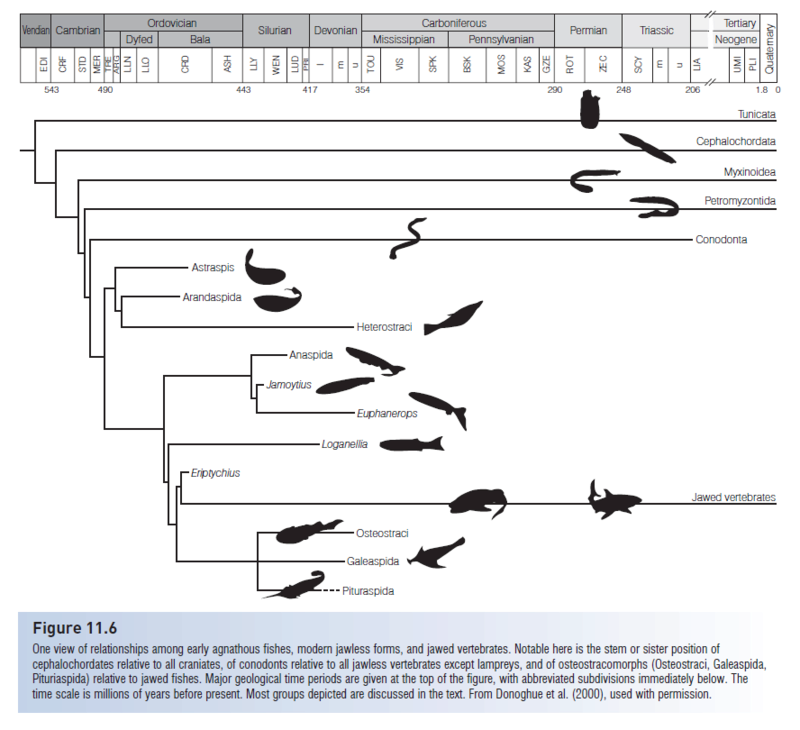
Pteraspidomorphi (or Diplorhina = “two nares”) derive their alternate name from impressions on the inside of the head plates indicating two separate olfactory bulbs in the brain. Pteraspidomorphs were jawless filter feeders in both marine and freshwater environments; they occurred from the Lower Silurian until the end of the Devonian. Three subclasses of pteraspidomorphs are recognized, the Astraspida Arandaspida, and Heterostraci. Primitive forms, such as the Ordovician Astraspis, Arandaspis, and arandaspid Sacabambaspis, had symmetrical tails, full body armor, and multiple branchial openings (Fig. 11.3).
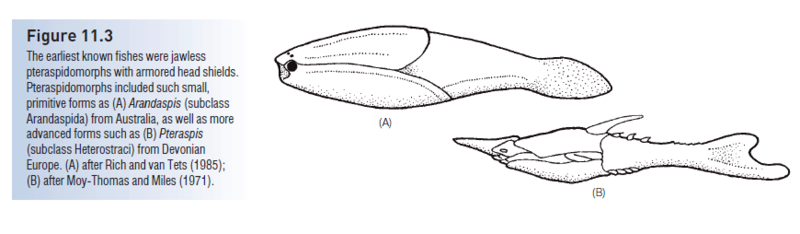
Heterostracans (“those with a different shell”) had dermal armor that extended from the head almost to the tail, necessitating swimming by lashing the tail back and forth, much like a tadpole. The tail in most forms was hypocercal, in that the notochord extended into the enlarged lower lobe of the tail. Their body form, armor, and tail morphology suggest that heterostracans plowed the bottom, pumping sediments into the ventral mouth, and filtering digestible material through the pharyngeal pouches. The armor is generally sutured and shows growth rings, indicating incremental growth. Early pteraspidiforms were small (c. 15 cm), but some heterostracans reached 1.5 m. Two orders, seven families, and more than 50 genera are recognized (see Denison 1970; Carroll 1988). Later heterostracans, such as the pteraspidiform Pteraspis from the Lower Devonian, had hypocercal tails, fused dorsal and ventral head plates, and single branchial openings (Fig. 11.3B). The Devonian also produced highly derived forms, such as the sawfishlike Doryaspis and the tube-snouted, blind Eglonaspis. Trends in the development of pteraspidiform lineages include the reduction of armor through fusion of plates, narrowing of the head shield, and development of lateral, presumably stabilizing, projections (cornua). These changes all suggest strong selection for increased mobility and maneuverability. While these anatomical changes were taking place, pteraspidiforms invaded freshwater habitats (Carroll 1988).
Chordate evolution and the three-phylum system 3
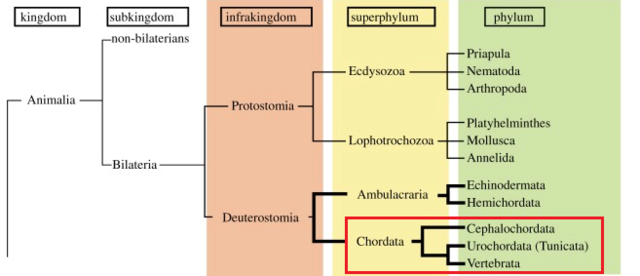
Traditional metazoan phylogeny classifies the Vertebrata as a subphylum of the phylum Chordata, together with two other subphyla, the Urochordata (Tunicata) and the Cephalochordata. The Chordata, together with the phyla Echinodermata and Hemichordata, comprise a major group, the Deuterostomia. Chordates invariably possess a notochord and a dorsal neural tube. Although the origin and evolution of chordates has been studied for more than a century, few authors have intimately discussed taxonomic ranking of the three chordate groups themselves. Accumulating evidence shows that echinoderms and hemichordates form a clade (the Ambulacraria), and that within the Chordata, cephalochordates diverged first, with tunicates and vertebrates forming a sister group. Chordates share tadpole-type larvae containing a notochord and hollow nerve cord, whereas ambulacrarians have dipleurula-type larvae containing a hydrocoel. We propose that an evolutionary occurrence of tadpole-type larvae is fundamental to understanding mechanisms of chordate origin. Protostomes have now been reclassified into two major taxa, the Ecdysozoa and Lophotrochozoa, whose developmental pathways are characterized by ecdysis and trochophore larvae, respectively. Consistent with this classification, the profound dipleurula versus tadpole larval differences merit a category higher than the phylum. Thus, it is recommended that the Ecdysozoa, Lophotrochozoa, Ambulacraria and Chordata be classified at the superphylum level, with the Chordata further subdivided into three phyla, on the basis of their distinctive characteristics.
Chordates consist of three distinct animal groups:
cephalochordates,
urochordates (tunicates)
vertebrates
Lamarck recognized Tunicata, namely animals enclosed with a tunic (tunica, in Latin, meaning garment). On the other hand, cephalochordates (lancelets) were first described in mid-to-late eighteenth century as molluscs. Although Yarrell had already noticed that lancelets have an axial rod, calling it ‘a lengthened internal vertebral column, although in a soft cartilaginous state’, it was Alexander Kowlevsky's discovery that both tunicates and lancelets possess notochords and dorsal neural tubes during embryogenesis, indicating that they are close relatives of vertebrates. He coined the name Chordonia for a hypothetical common ancestor of the Tunicata and the Vertebrata (including lancelets) by emphasizing the notochord as a significant diagnostic character shared by them. Later, Haeckel redefined Chordonia (i.e. Chordata) to include the Tunicata and the Vertebrata themselves. In London, Lankester gave subphylum status to the Urochordata, the Cephalochordata and the Craniata, altogether comprising the phylum Vertebrata. This constituted the first conception of the modern phylum Chordata.
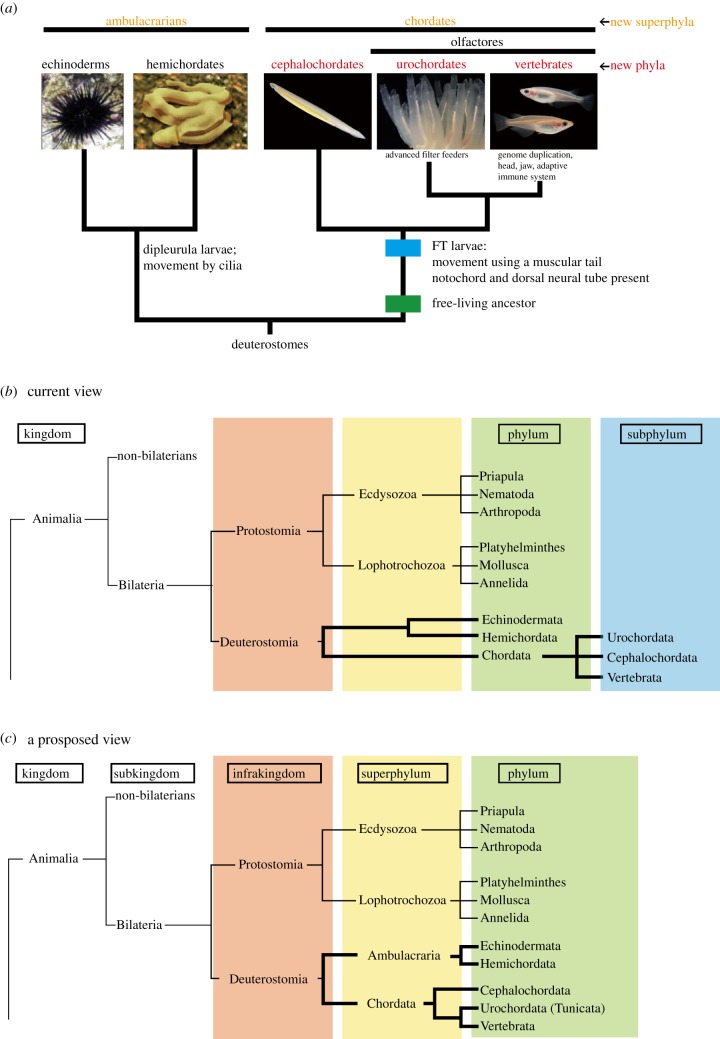
Phylogenic relationships of deuterostomes and evolution of chordates.
(a) Schematic representation of deuterostome groups and the evolution of chordates. Representative developmental events associated with the evolution of chordates are included. (b) A traditional and (c) the proposed view of chordate phylogeny with respect to their phylum relationship.
Bilaterians or triploblasts (metazoans composed of three germ layers: ectoderm, mesoderm and endoderm) are traditionally categorized into two major groups,
protostomes (in which the blastopore gives rise to the mouth) and
deuterostomes (in which the blastopore gives rise to the anus, and the mouth arises through secondary invagination of the stomodeum;
figure 1a), as first proposed by Grobben. Protostomes were further subdivided, mainly based on the mode of formation of the body cavity (or coelom), into acoelomates (with no distinct body cavity) such as platyhelminthes, pseudocoelomates (with a poorly developed body cavity) such as nematodes, and coelomates (with a distinct body cavity) such as annelids, molluscs and arthropods.
Molecular phylogeny, first based on comparison of 18S rDNA sequences and later protein-coding gene sequences, however, did not support this classification of protostomes, but instead suggested their division into two major groups, the Ecdysozoa (those exhibit moulting) and Lophotrochozoa (those having lophophores and trochophore larvae; sometimes called Spiralia; figure 1b). The former includes nematodes and arthropods, the latter annelids, molluscs and platyhelminths. The mode of body cavity formation therefore is not critical to protostome phylogeny, but developmental modes such as moulting and spiral cleavage are fundamental to evolutionary scenarios. This Ecdysozoa–Lophotrochoza classification has been supported by other studies, including Hox gene clustering, although there are several groups of which the phylogenic positions are still enigmatic, such as the mesozoans and chaetognaths.
On the other hand, recent studies of deuterostome molecular phylogeny, nuclear and mitochondrial genomics, and evolutionary developmental biology, have unambiguously demonstrated that echinoderms and hemichordates form a clade, and that urochordates, cephalochordates and vertebrates form another distinct clade (figure 1b). The former is called the Ambulacraria, with similarities in coelomic systems and larvae, and the latter Chordata. In addition, within the chordate clade, cephalochordates diverged first, and urochordates and vertebrates form a sister group (sometimes called Olfactores, with similarities in extensive pharyngeal re-modification leading to the formation of new structures, which are not found in cephalochordates). This novel view of deuterostome taxonomy and phylogeny became the consensus view, as a great variety of data from different disciplines support arguments for them.
The Xenacoelomorpha is a newly recognized phylum some have assigned to the deuterostomes, but this group is not discussed here because its phylogenetic position is still unstable.
1. https://en.wikipedia.org/wiki/Evolution_of_fish
2. THE DIVERSITY OF FISHES Biology, Evolution, and Ecology page 169
3. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4211455/
4. http://palaeos.com/vertebrates/vertebrata/craniata.html#Myllokunmingia
Further links:
[url=Vertebrate Paleobiology]https://www.geol.umd.edu/~jmerck/geol431/lectures/08osteichthyes.html[/url]
http://reasonandscience.heavenforum.org/t2482-evolution-of-fish
“As we move back down our evolutionary ladder, jaw structure becomes more instead of less complex, and in fishes the jaws are very elaborate indeed” (Maisey, 2000, p. 61).
Here we enter a strange and poorly known area of phylospace, one in which both timing and phylogeny are poorly understood. Somewhere between 750 and 550 million years ago, a new line of chordates evolved. These were characterised by a relatively concentrated set of sensory neurons at the anterior end and a particularly pronounced notochord extending beyond the anterior and posterior ends of the torso. In other words, they had the beginnings of a head and a tail. In addition, probably as a later development, the myomeres, the charcateristic bands of muscle along the torso, were reorganized in dorsal and ventral halves, separated by a septum, and acquired a distinctive 'V'- or 'W'-shape. These folks were our ancestors among chordate-kind. 4
The last common ancestor of hagfish and you, with all of its descendants, is referred to as Craniata. Craniates are animals with well-developed heads and a skull of sorts. A very early craniate, Myllokunmingia from the Cambrian of China, is badly illustrated in the figure at right. A rather better illustration of a possibly related and similar form is shown here. Since there is no really good reason to suppose that Haikouella belonged to this clade, we treat as a sort of "craniatomorph," but not a craniate.
From the first craniates evolved a bewildering variety of creatures. At least they have bewildered systematists for over a century. The first step seems relatively clear. The most primitive craniates we know are the living Myxinoidea (hagfishes): eel-like, rather assymmetric creatures with a strong propensity to flood their immediate neighborhood with slime at the slightest provocation. All other known craniates are Vertebrates. ATW020417.
There are three subphylums in chordata
Urochordata (tunicates)
Cephalochordata (lancelets)
Vertebrata
The First Vertebrates, Jawless Fishes, the Agnathans
Vertebrates arose over 500 million years ago (MYA) but traces of their appearance only occur during the Ordovician period about 460 MYA. These primeval, small fish-like vertebrates are popularly known as the ostracoderms and during the 100 million years of their existence they were comprised of about 600 species. Ostracoderms are especially important in the history and evolution of vertebrates. As undoubted vertebrates they possessed a backbone, were bilaterally symmetrical and had a nervous system divided into brain and spinal cord (partly enclosed within the backbone). They are also characterized by the possession of no more than two pairs of limbs and muscular system consisting primarily of bilaterally paired masses and a well-developed coelom, which contained the organs. In appearance, ostracoderms were dorsoventrally flattened and, quite extraordinarily, they lacked jaws, a condition so important that the classification of modern vertebrates is recognized by two major groups, the
agnathans (without jaws) and the
gnathostomes (literally, jaw-mouths)
Ostracoderms are now regarded as an artificial designation that includes perhaps four distinct superclasses of jawless craniate fishes, the Pteraspidomorphi, Anaspida, Thelodonti, and Osteostrachomorphi. Despite an apparently inauspicious beginning, these simple animals would be responsible for two events of major importance in the history of the vertebrates—the evolution of a tough outer protective layer, the integument, and the evolution of bone. Remarkably, in the beginning the histories of these seemingly disparate structures were intrinsically linked. Ostracoderms possessed an external bony head shield or armor but its internal skeleton was probably not ossified to any great extent. Debate over the origin of the vertebrate skeleton has revolved around the question of which group was the first to exhibit evidence of skeletonization and mineralization. The vertebrate skeleton is comprised of at least two distinct skeletal systems: the dermoskeleton (arising from the dermis; widely perceived to encompass teeth, scales, fin spines, etc.) and the endoskeleton (the braincase, branchial skeleton, axial, and appendicular ). It has been argued that although elements of the skeleton can be interchangeably derived from either system, the two systems have remained distinct throughout vertebrate phylogeny (Donoghue and Sansom 2002), of which more will be said later (Fig. 2.2).
Ostracoderms possessed an external bony head shield or armor but its internal skeleton was probably not ossified to any great extent. Debate over the origin of the vertebrate skeleton has revolved around the question of which group was the first to exhibit evidence of skeletonization and mineralization. The vertebrate skeleton is comprised of at least two distinct skeletal systems:
the dermoskeleton (arising from the dermis; widely perceived to encompass teeth, scales, fin spines, etc.) and the
endoskeleton (the braincase, branchial skeleton, axial, and appendicular skeletons).
It has been argued that although elements of the skeleton can be interchangeably derived from either system, the two systems have remained distinct throughout vertebrate phylogeny (Donoghue and Sansom 2002) (Fig. 2.2).
The evolution of fish began about 530 million years ago during the Cambrian explosion. According to the theory of evolution, life began in the seas, and the first advanced vertebrate animals were fish. Again according to the theory, these fish began to move toward dry land and in some way, came to use feet instead of fins and lungs instead of gills! Many books on evolution never consider the how of this major claim, whose baselessness is glossed over in most scientific textbooks in some summary like ". . . and living things moved from the water to dry land."
“A history of fishes” 2
Fishes were the first vertebrates. Understanding the
The focus is on fishes that lived during the Paleozoic and Mesozoic eras, and on modifications that occurred during the evolution of different, major extinct groups, leading to the dominant bony and cartilaginous fishes of today. We deal first with jawless fishes, then with ancestors of modern bony fishes because these occur earlier in the fossil record, and finally with the cartilaginous sharks, skates, rays, and chimaeras. This presentation focuses on extinct rather than extant fishes, recognizing that the distinctionb is artificial, that many lineages arose hundreds of millions of years ago and still have modern, living representatives, and that direct ancestors of some extant forms arose before other groups that have since gone extinct . We follow the basic organization of Nelson (2006) because of its synthetic and broad approach, recognizing that Nelson’s conclusions are one of many alternative interpretations of the literature.
The very first fishlike vertebrates undoubtedly evolved from invertebrates, perhaps a cephalochordate ( how do they know this is in sequence they admit that : ). However, the first “fishes” left no fossil record and their form and relationships remain a mystery. By the time fishlike fossils appear in Early Cambrian deposits, roughly 530 million years before present (Fig. 11.1), complex tissue types had evolved, including filamentous gills, V-shaped myomeres, and a distinct dorsal fin. New discoveries are made almost annually, but the currently recognized oldest species, Myllokunmingia fengjiaoa, was found in the Chengjiang geological formation of Yunnan Province in southwestern China. Myllokunmingia was 3– 4 cm long and is thought to be allied with (is a sister group to) ancestors of modern lampreys, although agreement is far from universal (Xian-guang et al. 2002;
Shu et al. 2003).
Lower Cambrian vertebrates from south China
NATURE | VOL 402 | 4 NOVEMBER 1999 |
The first fossil chordates are found in deposits from the Cambrian period (545±490 million years ago), but their earliest record is exceptionally sporadic and is often controversial.
Chordate is an animal belonging to the phylum Chordata; they possess a notochord, a hollow dorsal nerve cord, pharyngeal slits, an endostyle, and a post-anal tail, for at least some period of their life cycle. Chordates are deuterostomes, as during the embryo development stage the anus forms before the mouth. They are also bilaterally symmetric coelomates. In the case of vertebrate chordates, the notochord is usually replaced by a vertebral column during development, and they may have body plans organized via segmentation.
Accordingly, it has been diffcult to construct a coherent phylogenetic synthesis for the basal chordates. Until now, the available soft-bodied remains have consisted almost entirely of cephalochordate-like animals from Burgess Shale-type faunas. Definite examples of agnathan fish do not occur until the Lower Ordovician (,475 Myr BP), with a more questionable record extending into the Cambrian. The discovery of two distinct types of agnathan from the Lower Cambrian Chengjiang fossil-Lagerstaette is, therefore, a very significant extension of their range. One form is lampreylike, whereas the other is closer to the more primitive hagfish. These finds imply that the first agnathans may have evolved in the earliest Cambrian, with the chordates arising from more primitive deuterostomes in Ediacaran times (latest Neoproterozoic,,555 Myr BP), if not earlier.
If modern cephalochordates such as the lancelets (Branchiostoma) are considered fishlike
– if not exactly fishes –then the ancestry of fishes can be traced farther back to the cephalochordate-like yunnanozoans (Haikouella and Yunnanozoon) from the Lower Cambrian, or to the muchheralded Pikaia with its dorsal nerve cord and notochord, from the Middle Cambrian Burgess Shale of British Columbia. Clearly recognizable fish specimens, such as the arandaspid pteraspidomorph Sacabambaspis janvieri from Bolivia, appear later, dating to 470 mybp (Gagnier et al. 1986; Gagnier 1989). This and related jawless, finless forms inhabited shallow seas or estuarine habitats in tropical and subtropical regions of the Gondwanan and Laurasian supercontinents. Their innovations include true bone (probably evolved independently in several ancestral groups) and a muscular feeding pump. The former adaptation, which existed only as an external covering, would have provided protection from predators to which softer ancestors were more vulnerable, as well as serving as a metabolic reserve for calcium and phosphate and an insulator of electrosensory organs (Northcutt & Gans 1983; Carroll 1988). A muscular feeding pump would have been more efficient for moving food-bearing water through a filtration mechanism than was the ciliary feeding mechanism of protochordates. Another major advance over the cephalochordates that preceded them was that, although lacking jaws, the early fossilized fishes were craniates. They had a head region containing a brain with specialized sensory capsules and cranial nerves, all contained in a protective skeletal braincase (Maisey 1996).
Subphylum Craniata
Vertebrate craniates possess, among other features, a dermal skeleton and neural crest, the latter describing regions of the developing nerve cord that are precursors to gill arches, pigment cells, connective tissue, and bone. Within the vertebrates are seven superclasses of fishes, five of which are extinct.
†Pteraspidomorphi
Pteraspidomorphi (or Diplorhina = “two nares”) derive their alternate name from impressions on the inside of the head plates indicating two separate olfactory bulbs in the brain. Pteraspidomorphs were jawless filter feeders in both marine and freshwater environments; they occurred from the Lower Silurian until the end of the Devonian. Three subclasses of pteraspidomorphs are recognized, the Astraspida Arandaspida, and Heterostraci. Primitive forms, such as the Ordovician Astraspis, Arandaspis, and arandaspid Sacabambaspis, had symmetrical tails, full body armor, and multiple branchial openings (Fig. 11.3).
Heterostracans (“those with a different shell”) had dermal armor that extended from the head almost to the tail, necessitating swimming by lashing the tail back and forth, much like a tadpole. The tail in most forms was hypocercal, in that the notochord extended into the enlarged lower lobe of the tail. Their body form, armor, and tail morphology suggest that heterostracans plowed the bottom, pumping sediments into the ventral mouth, and filtering digestible material through the pharyngeal pouches. The armor is generally sutured and shows growth rings, indicating incremental growth. Early pteraspidiforms were small (c. 15 cm), but some heterostracans reached 1.5 m. Two orders, seven families, and more than 50 genera are recognized (see Denison 1970; Carroll 1988). Later heterostracans, such as the pteraspidiform Pteraspis from the Lower Devonian, had hypocercal tails, fused dorsal and ventral head plates, and single branchial openings (Fig. 11.3B). The Devonian also produced highly derived forms, such as the sawfishlike Doryaspis and the tube-snouted, blind Eglonaspis. Trends in the development of pteraspidiform lineages include the reduction of armor through fusion of plates, narrowing of the head shield, and development of lateral, presumably stabilizing, projections (cornua). These changes all suggest strong selection for increased mobility and maneuverability. While these anatomical changes were taking place, pteraspidiforms invaded freshwater habitats (Carroll 1988).
Chordate evolution and the three-phylum system 3
Traditional metazoan phylogeny classifies the Vertebrata as a subphylum of the phylum Chordata, together with two other subphyla, the Urochordata (Tunicata) and the Cephalochordata. The Chordata, together with the phyla Echinodermata and Hemichordata, comprise a major group, the Deuterostomia. Chordates invariably possess a notochord and a dorsal neural tube. Although the origin and evolution of chordates has been studied for more than a century, few authors have intimately discussed taxonomic ranking of the three chordate groups themselves. Accumulating evidence shows that echinoderms and hemichordates form a clade (the Ambulacraria), and that within the Chordata, cephalochordates diverged first, with tunicates and vertebrates forming a sister group. Chordates share tadpole-type larvae containing a notochord and hollow nerve cord, whereas ambulacrarians have dipleurula-type larvae containing a hydrocoel. We propose that an evolutionary occurrence of tadpole-type larvae is fundamental to understanding mechanisms of chordate origin. Protostomes have now been reclassified into two major taxa, the Ecdysozoa and Lophotrochozoa, whose developmental pathways are characterized by ecdysis and trochophore larvae, respectively. Consistent with this classification, the profound dipleurula versus tadpole larval differences merit a category higher than the phylum. Thus, it is recommended that the Ecdysozoa, Lophotrochozoa, Ambulacraria and Chordata be classified at the superphylum level, with the Chordata further subdivided into three phyla, on the basis of their distinctive characteristics.
Chordates consist of three distinct animal groups:
cephalochordates,
urochordates (tunicates)
vertebrates
Lamarck recognized Tunicata, namely animals enclosed with a tunic (tunica, in Latin, meaning garment). On the other hand, cephalochordates (lancelets) were first described in mid-to-late eighteenth century as molluscs. Although Yarrell had already noticed that lancelets have an axial rod, calling it ‘a lengthened internal vertebral column, although in a soft cartilaginous state’, it was Alexander Kowlevsky's discovery that both tunicates and lancelets possess notochords and dorsal neural tubes during embryogenesis, indicating that they are close relatives of vertebrates. He coined the name Chordonia for a hypothetical common ancestor of the Tunicata and the Vertebrata (including lancelets) by emphasizing the notochord as a significant diagnostic character shared by them. Later, Haeckel redefined Chordonia (i.e. Chordata) to include the Tunicata and the Vertebrata themselves. In London, Lankester gave subphylum status to the Urochordata, the Cephalochordata and the Craniata, altogether comprising the phylum Vertebrata. This constituted the first conception of the modern phylum Chordata.
Phylogenic relationships of deuterostomes and evolution of chordates.
(a) Schematic representation of deuterostome groups and the evolution of chordates. Representative developmental events associated with the evolution of chordates are included. (b) A traditional and (c) the proposed view of chordate phylogeny with respect to their phylum relationship.
Bilaterians or triploblasts (metazoans composed of three germ layers: ectoderm, mesoderm and endoderm) are traditionally categorized into two major groups,
protostomes (in which the blastopore gives rise to the mouth) and
deuterostomes (in which the blastopore gives rise to the anus, and the mouth arises through secondary invagination of the stomodeum;
figure 1a), as first proposed by Grobben. Protostomes were further subdivided, mainly based on the mode of formation of the body cavity (or coelom), into acoelomates (with no distinct body cavity) such as platyhelminthes, pseudocoelomates (with a poorly developed body cavity) such as nematodes, and coelomates (with a distinct body cavity) such as annelids, molluscs and arthropods.
Molecular phylogeny, first based on comparison of 18S rDNA sequences and later protein-coding gene sequences, however, did not support this classification of protostomes, but instead suggested their division into two major groups, the Ecdysozoa (those exhibit moulting) and Lophotrochozoa (those having lophophores and trochophore larvae; sometimes called Spiralia; figure 1b). The former includes nematodes and arthropods, the latter annelids, molluscs and platyhelminths. The mode of body cavity formation therefore is not critical to protostome phylogeny, but developmental modes such as moulting and spiral cleavage are fundamental to evolutionary scenarios. This Ecdysozoa–Lophotrochoza classification has been supported by other studies, including Hox gene clustering, although there are several groups of which the phylogenic positions are still enigmatic, such as the mesozoans and chaetognaths.
On the other hand, recent studies of deuterostome molecular phylogeny, nuclear and mitochondrial genomics, and evolutionary developmental biology, have unambiguously demonstrated that echinoderms and hemichordates form a clade, and that urochordates, cephalochordates and vertebrates form another distinct clade (figure 1b). The former is called the Ambulacraria, with similarities in coelomic systems and larvae, and the latter Chordata. In addition, within the chordate clade, cephalochordates diverged first, and urochordates and vertebrates form a sister group (sometimes called Olfactores, with similarities in extensive pharyngeal re-modification leading to the formation of new structures, which are not found in cephalochordates). This novel view of deuterostome taxonomy and phylogeny became the consensus view, as a great variety of data from different disciplines support arguments for them.
The Xenacoelomorpha is a newly recognized phylum some have assigned to the deuterostomes, but this group is not discussed here because its phylogenetic position is still unstable.
1. https://en.wikipedia.org/wiki/Evolution_of_fish
2. THE DIVERSITY OF FISHES Biology, Evolution, and Ecology page 169
3. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4211455/
4. http://palaeos.com/vertebrates/vertebrata/craniata.html#Myllokunmingia
Further links:
[url=Vertebrate Paleobiology]https://www.geol.umd.edu/~jmerck/geol431/lectures/08osteichthyes.html[/url]
Last edited by Admin on Sat Aug 19, 2017 7:41 pm; edited 4 times in total