The recent groundbreaking scientific research which explains the real mechanisms of biodiversity
https://reasonandscience.catsboard.com/t2293-the-recent-groundbreaking-scientific-research-which-explains-the-real-mechanisms-of-biodiversity
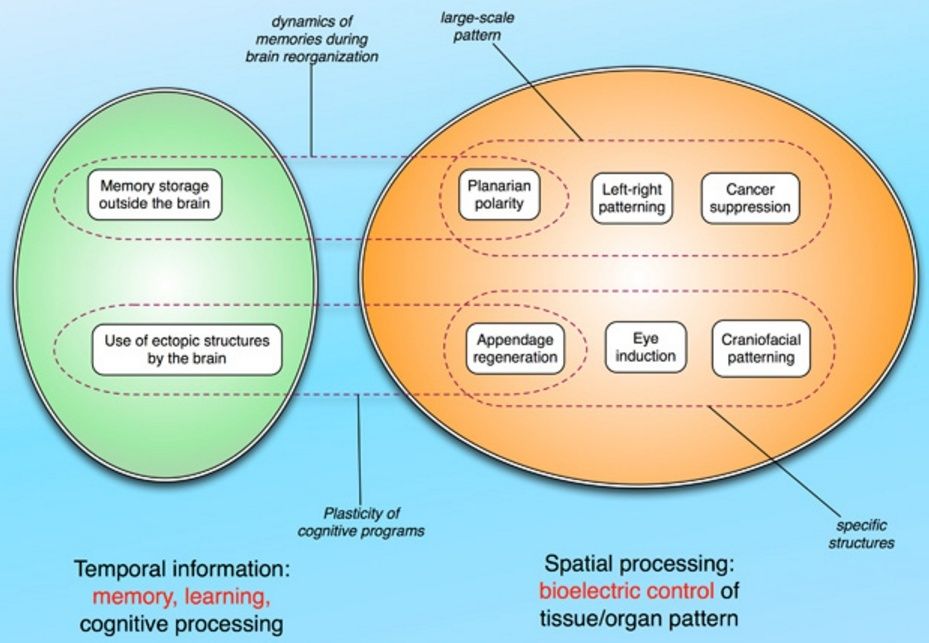
The assertion that evolution is a fact, is repeated like a mantra by proponents of evolution and naturalism, which try in that way to justify their unbelief in an intelligent creator. One of the most frequent claims is that microevolution and macroevolution are the same on a different timescale. And that there is no mechanism that prevents micro to become macro. The ones that are better informed, IMHO, know that the mechanism that provokes change and evolutionary novelties above the species level, that is, the change from bacteria do man, is UNKNOWN. In order to explain the origin of biodiversity, body shape, body plans, organ development, and ultimately, if the claim of macro-evolution from Luca to homo sapiens is true, we need first to understand how organs, limbs, and creatures arise, what mechanism determines what size and shape they should be when they are growing. What mechanism programs the cell to "know" how and where to form an organ? How does the creature know how to rebuild a severed limb to the correct size, shape, and orientation? Once this is elucidated, we can ask if the same mechanisms explain biodiversity. It seems that groundbreaking scientific research is starting to unravel this longstanding mystery, and it's far from being explained through neodarwiniam predictions and claims, but epigenetic mechanisms, which will be elucidated below.
A great deal is now known about morphogen gradients in the developing embryo—how cells know where to go and what types of cells to become. Recent research shows that electric potentials in nonbrain cells are a signal for creating patterns during the development and during the rebuilding of organs. This information of different field potentials surrounding individual cells can give information for the developing organ. Now, research from the laboratory of Dr. Michael Levin is demonstrating new ways that cells signal with electricity and the great importance of electrical properties for individual cells and tissues. He describes how electrical gradients and fields are critical in the 3D function and shape of cells and organs.
A prominent aspect of multi-cellular creatures is that they have organs of a particular size and shape. When regeneration occurs in reptiles the same exact shape is grown. Information for the cellular activity appears to exist in the space that will make up the specifically shaped organ. Levin notes that “cancer can be seen as an error of geometry, because tumor cells grow, migrate, and function without regard for the orderly structure within which they occur.”
Each cell has specific electrical gradients and properties that together form a large electric field of information. This field of information can show individual cells in the embryo how to behave. This is analogous to the fact that electrical flow between cells in the early embryo forms the basic network that is then built into a formal structure with elaborate chemical synapses. This, also, occurs during rebuilding of tissue. Somehow, the information of the electrical flow through the electric gap junction synapses determines the future structure. Electrical signals now appear to be critical in forming the shape of organs, the very function and identity of organs, and the creation of new limbs on animals that regenerate. In these animals, stem cell behavior is directed by currents created with potassium, sodium, chloride, and protons that affect the genetic networks of cells at a distance. Recent research shows that limbs can be influenced by proton and sodium alterations. New types of ion channels, pumps and electrical connections have now been found in a variety of different organs.
This is remarkable. If electric gap junction synapses determine cell shape, then macro change attributed commonly to macroevolution and structural biological novelties should also depend on these.
Electrical signaling is key for cells to properly interpret their environment, and when this process goes awry, the cells default to a cancer program. 4
While ion flows control cell-level behaviors such as migration, differentiation, and proliferation, bioelectric signals also function as master regulators of large-scale shape in many contexts: a simple signal can induce complex, highly orchestrated, self-limiting downstream morphogenetic cascades. For example, an unmodulated flux of protons can cause the formation of a complete tail of the right rise and tissue composition.
Our data suggest that the mechanism by which blastema cells polls the rest of the host (to determine where the wound is located and what other tissues already exist in the fragment and thus don't need to be recreated) is mediated by physiological signals passing through nerves and long-range gap junctional paths.
A significant component of morphogenetic cues are ionic in nature. Remarkably, however, this effect is non-local in nature - it is the transmembrane potential of other, quite distant cells that determines the metastasis-like effect.
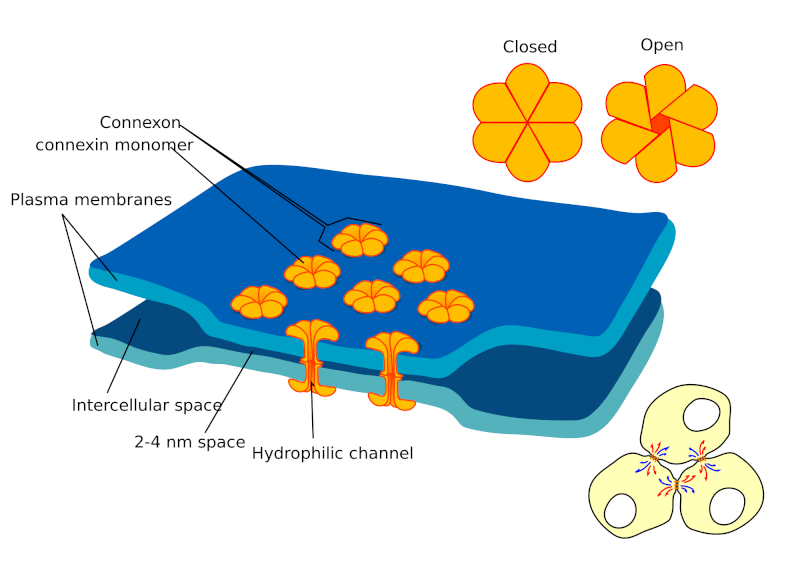
Many critical questions remain about how cellular polarity is synchronized and amplified across embryonic fields to allow cells to ascertain their position with respect to the midline. We identified a dependence of asymmetric gene expression on early communication between left and right sides in the chick and frog. For example, expression of left-sided markers depends on events occurring on the right side, during very early stages, suggesting that the two sides need to coordinate their decision with respect to the L-R identity of each. One mechanism for communicating between cells and tissues involves gap junctions: multimers of connexin proteins form channels between cells and pass small molecules, subject to complex regulation by various signals.
So it seems within gap junctions that action happens, but the action per se is REGULATION AND COMMUNICATION THROUGH VARIOUS SIGNALS. THATS THE KEY.
We showed that gap junction are crucially involved in L-R patterning in early embryos of Xenopus and chick. gap junctions are a bioelectric patterning element that sets up domains of isopotential cell fields during morphogenesis.
Serotonin signaling is used in information exchange between cells in processes such as L-R patterning and control of timing and cell movement during gastrulation. We have shown that serotonin is utilized by both chick and frog embryos, at very early stages, as a small molecule signal which is transported in a left-right gradient and regulates the development of laterality. Indeed, we now know that the early frog embryo is literally an electrophoresis chamber, which uses voltage potentials to generate consistently biased left-right gradients in serotonin in an epigenetic process not dependent on zygotic gene expression. We have modeled this process quantitatively, and characterized novel intracellular serotonin-binding proteins which directly activate asymmetric gene expression after their rightward movement, linking an early biophysical process to transcriptional regulation via chromatin modification pathways. Serotonin is also a key mediator for bioelectric control of neuronal outgrowth from transplants.
Electrical Fields Guiding 3D Shape of Cells and Organs
How does the cell know what size and shape it should be? Many cells alter their shape to provide different functions, like microglia. Even more complex is the question as to how organs, limbs, and creatures know what size and shape they should be when they are growing. How do the cells know how and where to form an organ? How does the creature know how to rebuild a severed limb to the correct size, shape, and orientation?
There are thousands of these same questions, including how astrocytes and neurons know the exact networks they should form. How immune cells know how to travel through very complex differing 3D environments? An equally difficult question has been addressed in previous posts as to how the
cell knows exactly what shape a coded sequence in a protein will take. Both cells and microbes alter the codes of their protein toxins that require extremely detailed and accurate shapes. In fact, modern science cannot calculate what shape a 400 amino acid coded sequence will be when it folds into a protein. It would take all the supercomputers together two thousand years to calculate the folding of one average protein. Yet, proteins assemble into the exact shape in a millisecond, helped by very complex chaperone molecules. Cells routinely edit their messenger RNA with alternative splicing to make a whole variety of shapes. How do they know how to do this?
Research shows that: If a cell of a two cell embryo is removed it still becomes the full creature.
Starved flatworms shrink keeping perfect proportions among the organs
Planaria can reproduce the entire body from a small piece
Amphibians can grow a perfectly proportioned limb
Many structures need nerve-mediated information in order to maintain shape and function
For all of these questions, it is hard to imagine the information of the 3D shape being inside one cell, or even a group of cells. It seems more likely that a field of information somehow guides these processes. Is this evidence for an electrical field of information forming 3D shapes of cells, organs and creatures, also, related to the influence of mind?
Previous posts have demonstrated the critical part that electrical synapses play in forming the structure of the neuronal network that uses chemical synapses. Also, a previous post showed that field potentials in and around brain cells, while poorly understood, are important for specific functions. Multiple posts have documented the elaborate communication that occurs between cells through signaling including neurotransmitters, brain factors, cytokines and hormones.
A significant amount of research shows the importance of various gradients in the space between cells for communication. This was noted in the posts on the movement of platelets that attract and signal to other immune cells. Another post showed the elaborate travels of leukocytes. A great deal is now known about morphogen gradients in the developing embryo—how cells know where to go and what types of cells to become.
Recent research shows that electric potentials in non brain cells are a signal for creating patterns during development and during re building of organs. This information of different field potentials surrounding individual cells can give information for the developing organ. Now, research from the laboratory of Dr. Michael Levin is demonstrating new ways that cells signal with electricity and the great importance of electrical properties for individual cells and tissues. He describes how electrical gradients and fields are critical in the 3D function and shape of cells and organs. Professor Levin notes that cancer cells use electric gradients for many purposes. In fact, aberrations in electrical processes can lead to cancer. Differences in electrical potential in cells are important in determining whether a cancer metastasizes and electrical states in one cell can have effects on distant cells, triggering metastasis. These electrical potentials in one cell can signal other cells that trigger genetic networks and epigenetic effects.

From Magnus Manske
The electric state of a cell is determined by many complex factors. One is the opening and closing of ion channel proteins, which control the movement of charged particles (ions) across the cell membrane. Changes of the cells’ electrical potential via the activity of ion channels are shown to be able to suppress or trigger cancer. Another factor is electrical synapses that transfer electricity from one cell to another.
Different electrical properties of cells affect what type of cells are made from stem cells, how much reproduction goes on, where cells travel to and their shape and orientation. Electrical gradients and fields certainly affect travelling immune cells, stem cells, and all brain cells. They also exert profound effects on other somatic cell types during embryonic development and regenerative repair. Electrical properties are simultaneous with chemical metabolism using genetic network processing. They are critical for healing wounds and responding to infections. They determine the symmetry and shape of organs.
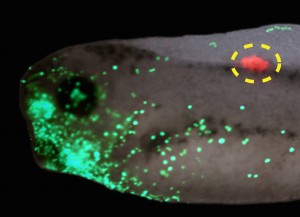
Differential ion channel activity in the body sets up specific patterns of voltage potential throughout tissues. During development, these electrical properties of cells determine what types of cells they become. For example, two specific groups of cells in the embryo have increased electrical potential and these become the two eyes. Research shows that by changing the electrical potential of cells at a critical time when eyes are being made, eyes can be induced to form elsewhere, effectively reprogramming other cell types into a complete visual organ.
While electrical activity can drive downstream genetic changes, just changing the electrical gradient can dramatically change the cell’s behavior. After an amputation, controlling the voltage potentials of wound cells causes the regrowth of a limb. The specification of the details appears to exist in the electrical field. At specific regions of the animal’s body, either a tail or limb grow due to the electrical signaling, which kickstarts a “build whatever goes here” program.
Electrical fields that are being discovered with multiple influences could contain this 3D information. Altering the electric field alters the information and the biological results.
Quite a number of previous posts reveal the elaborate signaling of microbes and immune cells such as T cells and platelets. The critical intestinal epithelial cells and skin cells are involved in very elaborate signaling between large numbers of microbes on one side and many immune cells on the others. Of course, brains are known to use elaborate signaling.
A previous post noted that cancer is like a microbe colony in that they work together for their ends. This view of cancer is that many cells in an environment trigger the actions of the community of cancer cells. Another view of cancer is as a defection of individual cells from the goals of the organism; this can occur by cells’ being isolated from, or mis-interpreting, the patterning cues that normally orchestrate cell behavior into complex anatomy. The new research by Levin adds a very large new dimension in the way electrical signals between cells might be critical in this cancer development.
A prominent aspect of multi cellular creatures is that they have organs of a particular size and shape. When regeneration occurs in reptiles the same exact shape is grown. Information for the cellular activity appears to exist in the space that will make up the specifically shaped organ. Levin notes that “cancer can be seen as an error of geometry, because tumor cells grow, migrate, and function without regard for the orderly structure within which they occur.”
This can occur through each cell having specific electrical gradients and properties that together form a large electric field of information. This field of information can show individual cells in the embryo how to behave. This is analogous to the fact that electrical flow between cells in the early embryo (see post on electrical synapses) forms the basic network that is then built into a formal structure with elaborate chemical synapses. This, also, occurs during rebuilding of tissue. Somehow, the information of the electrical flow through the electric gap junction synapses determines the future structure.
Electrical signals now appear to be critical in forming the shape of organs, the very function and identity of organs, and the creation of new limbs on animals that regenerate. In these animals, stem cell behavior is directed by currents created with potassium, sodium, chloride, and protons that affect the genetic networks of cells at a distance. Recent research shows that limbs can be influenced by proton and sodium alterations. New types of ion channels, pumps and electrical connections have now been found in a variety of different organs.
This is remarkable. If electric gap junction synapses determine cell shape, then macro change attributed commonly to macroevolution and structural biological novelties should also depend on these.
Cells not in the brain, also, use ion channels that create an electrical potential between the inside and outside of the cell. These do not send a signal down a long axon wire as neurons do, but the electrical potentials are critical for the cell’s behavior. In a previous post, calcium cyclic signaling was noted to be the crucial signal between plants and microbes when forming nitrogen factories.
Whole tissues and sheets of cells, also, have flows of electricity through electrical synapses called gap junctions (just like neurons). When the skin is broken the electrical gradients and fields are disrupted and these are a signal for immune cells to repair the tissue. An even larger field exists over an entire organ, and this field has crucial information regarding the shape and function of the organ.
It is very difficult to study, but even inside the cell, many membranes exist for the nucleus, the mitochondria and other organelles such as the critical information center of the endoplasmic reticulum. Electrical potentials and information fields exist in these, also.
Cancer cells’ behavior is very tied to electrical signals. They use them as clues to avenues of travel. Electrical fields determine shape changes in the cells. The electrical potential, also, affects what type of cell is produced from stem cells.
Cancer cells have altered electrical activity and use different ion channels and pumps. They have different transporter molecules that affect how the ions travel and, therefore, the electrical gradients. It now appears that these alterations in ion channels are signatures of different cancers and are critical to their formation. Many differences in the effects of cytokines and neurotransmitter signals, also, appear to be tied to these electrical differences. These ion channels influence all aspects of metastasis. In fact, oncogenes (special genes that cause a cell to become cancerous) are related to various different ion channels.
Research in cancer has been increasingly difficult because of the many different mutations that are found in types of cancers, and, more recently, in individuals. The studies have shown that there is a wide range of different mutations that don’t fall into any noticeable patterns.
Dr. Levin’s work points in a different direction entirely. He notes that the gradients and fields of electrical information may be a more important way to understand this process. It is the very space that the cells are living in that appears to be aberrant and causing the cancers. Electrical signaling is key for cells to properly interpret their environment, and when this process goes awry, the cells default to a cancer program.
Studies show that cancer cells can be differentiated from other cells by their electrical properties. The overall electrical status of the cancer cell is determined by the sum of all the ions and pumps. There are a vast amount of different possible genetic scenarios in this basic measurement of electrical status.
But, in fact, the simple electrical status appears to identify cancer cells. Dr. Levin likens this to “pressure” in physics – a group property that can be implemented with a wide variety of underlying molecular details (e.g., different ion channel genes). He notes that the statistical electrical study of cancer electric gradients might be the most information. He notes that the extremely complex analysis of all of the ion channels might not even reveal the critical information (this type of research is currently not even feasible).
Just analyzing messenger RNA and specific proteins made will not determine all the factors altering the global electrical fields because electrical state is a function of the 3D open/closed states of the channels, not merely their presence at the mRNA and protein levels.
The large electrical fields themselves appear to create the regulation that occurs. A study showed that by increasing the electric potential in cancer cells, metastatic activity can be suppressed, despite specific proteins being present that are known to be involved in metastatic activity.
Information Fields and Geometry
All of the signals that affect a cell, together, are called the morphogenic field. It includes all of the instructions that come from all secreted cellular signals, including cytokines and the effects of all the molecules in the extra cellular space (see post on Extracellular Space and Neuroplasticity). This includes many gradients that are used by immune cells to gather information and attract other cells. An unusual feature recently discovered as a cause of cancer is special proteins that produce cells in particular orientations—called planar cell polarity. When this polarity is altered it can cause cancer.
Electrical gradients are now shown to be critical as well. When the usual gradient or field information is somehow altered it can cause cancer. Each embryo has its own innate field. It has been observed that cells in the midst of a live organ are not as susceptible to the same chemical influences to become cancerous. They are much more able to become cancer when isolated from the influences of other cells. When the region’s electrical properties are different, they are less likely to become cancerous.
Signals from neurons are, also, a factor in causing cancer. When nerves are cut, then cancers occur more frequently. These neuron signals appear to be contributing to the field information in local regions.
A striking example of the field is when shunts are needed between the abdomen and the blood. These shunts send large amounts of cancer cells to many regions, but they do not become metastatic in some areas, but do in others. Another is when the shape of a region is altered by surgery it is more likely to become cancerous, possibly by altering the field.
Cancer cells turn away from the community aspect of the entire organism and become involved in individual behavior and their own new community (see post on Cancer – The Emperor of Cells.) The cancer cells either aren’t given appropriate information in the field about the creature, can’t extract the information or don’t care anymore. Cancers form their own organs and fields. The cancer cells, also, form their own information fields and communicate and cooperate using them. They reproduce and evolve to be stronger as a community. They signal each other to fight microbes and immune attacks. Cancer cells eliminate communication to the body and tissues of the body. But, they maintain the same junctions and communications among this smaller community.
The study of electric fields brings up the problem of orders of magnitude of effect and the question of emergent properties. While at the level of genetic network activity, each cell in the field has definite behaviors—they are all influenced by the overall information fields. All levels are at play—the genetic, epigenetic, individual cells and networks of cells.
Having information in morphogenic fields including electrical gradients is similar to the notion that mental events occur in neuronal electric networks. Dr. Levin speculates that the same way that information is contained in neural networks, information could be maintained in electrical fields of cells, organs and creatures.
All of this, raises the question, of course, about its relation to cellular intelligence in particular and mind in nature in general.
The genetic networks are very critical in electric signaling because cells use protein ion channels imbedded in the membrane. These proteins affect distant actions and are very involved in creating the overall gradients.
In one of many circular genetic processes, the actions of the genetic machinery are controlled by electrical gradients and the elaborate signaling of the genetic networks create the electrical gradient.
Nices Catch22 situation.
Patterning occurs by a continuous interplay of genetics and physics.
This conundrum indicates that there is a more global regulation of the entire process. In fact, research shows that the electrical influences cannot be reduced to simple chemistry inside of a cell. Electrical gradients of individual cells and many cells contribute to the creation of the 3D shapes and functions of organs and creatures. Examples of this include the creation of the embryonic eye, the head, and the regeneration of limbs exactly to size. For movement of cells, when electrical and chemical gradients are in competition, the electrical gradient wins.
The example of the eye is instructive. Dr. Levin notes that it shows the electrical information is more crucial. It is the electrical properties of the regions forming the two eyes that determine where the eyes will go. When this electrical property is altered, the eye forms in different places. This effect cannot be replicated outside of the head by any of the known “master” eye genes.
How does a cell first create a shape of an organ in the fetus and then maintain it throughout life, with many new cells being generated? Signals can let a cell know about position, size and polarity and orientation. It is either cell-to-cell communication, which is basically the current research goal, or a pre formed field or organizing template. The 3D process in the fetus is called morphogenesis. Each cell in the embryo has to know its position relative to others and differentiates into the appropriate type of cell for that position.
Field structures in physics are similar to the concept of morphogenetic field. A field includes information at each location. It includes regulation at a distance. The gradient is a concept already used in studies of development. Current research shows that electrical potentials and gradients in the space between cells are an important source of information for cells, and control the movement of chemical signals such as serotonin and butyrate.
Electrical Fields Guiding 3D Shape of Cells and Organs
While both chemical and electrical factors are clearly relevant to determining the three dimensional function and shapes of cells, organs and creatures, it appears that an information field of some kind would be necessary for the level of information needed. Recent research shows the importance of the electrical potentials, gradients and fields.
Among theories of what mind could be (see post), information fields have been proposed. Perhaps this current research furthers such a view.
1) http://reasonandscience.heavenforum.org/t1826-how-does-evolution-supposedly-work
2) http://reasonandscience.heavenforum.org/t2279-what-prevents-the-transition-from-micro-to-macro-evolution
3) http://jonlieffmd.com/blog/electrical-fields-guiding-3d-shape-of-cells-and-organs
4) http://www.drmichaellevin.org/research.html
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3413735/
https://reasonandscience.catsboard.com/t2293-the-recent-groundbreaking-scientific-research-which-explains-the-real-mechanisms-of-biodiversity
The assertion that evolution is a fact, is repeated like a mantra by proponents of evolution and naturalism, which try in that way to justify their unbelief in an intelligent creator. One of the most frequent claims is that microevolution and macroevolution are the same on a different timescale. And that there is no mechanism that prevents micro to become macro. The ones that are better informed, IMHO, know that the mechanism that provokes change and evolutionary novelties above the species level, that is, the change from bacteria do man, is UNKNOWN. In order to explain the origin of biodiversity, body shape, body plans, organ development, and ultimately, if the claim of macro-evolution from Luca to homo sapiens is true, we need first to understand how organs, limbs, and creatures arise, what mechanism determines what size and shape they should be when they are growing. What mechanism programs the cell to "know" how and where to form an organ? How does the creature know how to rebuild a severed limb to the correct size, shape, and orientation? Once this is elucidated, we can ask if the same mechanisms explain biodiversity. It seems that groundbreaking scientific research is starting to unravel this longstanding mystery, and it's far from being explained through neodarwiniam predictions and claims, but epigenetic mechanisms, which will be elucidated below.
A great deal is now known about morphogen gradients in the developing embryo—how cells know where to go and what types of cells to become. Recent research shows that electric potentials in nonbrain cells are a signal for creating patterns during the development and during the rebuilding of organs. This information of different field potentials surrounding individual cells can give information for the developing organ. Now, research from the laboratory of Dr. Michael Levin is demonstrating new ways that cells signal with electricity and the great importance of electrical properties for individual cells and tissues. He describes how electrical gradients and fields are critical in the 3D function and shape of cells and organs.
A prominent aspect of multi-cellular creatures is that they have organs of a particular size and shape. When regeneration occurs in reptiles the same exact shape is grown. Information for the cellular activity appears to exist in the space that will make up the specifically shaped organ. Levin notes that “cancer can be seen as an error of geometry, because tumor cells grow, migrate, and function without regard for the orderly structure within which they occur.”
Each cell has specific electrical gradients and properties that together form a large electric field of information. This field of information can show individual cells in the embryo how to behave. This is analogous to the fact that electrical flow between cells in the early embryo forms the basic network that is then built into a formal structure with elaborate chemical synapses. This, also, occurs during rebuilding of tissue. Somehow, the information of the electrical flow through the electric gap junction synapses determines the future structure. Electrical signals now appear to be critical in forming the shape of organs, the very function and identity of organs, and the creation of new limbs on animals that regenerate. In these animals, stem cell behavior is directed by currents created with potassium, sodium, chloride, and protons that affect the genetic networks of cells at a distance. Recent research shows that limbs can be influenced by proton and sodium alterations. New types of ion channels, pumps and electrical connections have now been found in a variety of different organs.
This is remarkable. If electric gap junction synapses determine cell shape, then macro change attributed commonly to macroevolution and structural biological novelties should also depend on these.
Electrical signaling is key for cells to properly interpret their environment, and when this process goes awry, the cells default to a cancer program. 4
While ion flows control cell-level behaviors such as migration, differentiation, and proliferation, bioelectric signals also function as master regulators of large-scale shape in many contexts: a simple signal can induce complex, highly orchestrated, self-limiting downstream morphogenetic cascades. For example, an unmodulated flux of protons can cause the formation of a complete tail of the right rise and tissue composition.
Our data suggest that the mechanism by which blastema cells polls the rest of the host (to determine where the wound is located and what other tissues already exist in the fragment and thus don't need to be recreated) is mediated by physiological signals passing through nerves and long-range gap junctional paths.
A significant component of morphogenetic cues are ionic in nature. Remarkably, however, this effect is non-local in nature - it is the transmembrane potential of other, quite distant cells that determines the metastasis-like effect.
Many critical questions remain about how cellular polarity is synchronized and amplified across embryonic fields to allow cells to ascertain their position with respect to the midline. We identified a dependence of asymmetric gene expression on early communication between left and right sides in the chick and frog. For example, expression of left-sided markers depends on events occurring on the right side, during very early stages, suggesting that the two sides need to coordinate their decision with respect to the L-R identity of each. One mechanism for communicating between cells and tissues involves gap junctions: multimers of connexin proteins form channels between cells and pass small molecules, subject to complex regulation by various signals.
So it seems within gap junctions that action happens, but the action per se is REGULATION AND COMMUNICATION THROUGH VARIOUS SIGNALS. THATS THE KEY.
We showed that gap junction are crucially involved in L-R patterning in early embryos of Xenopus and chick. gap junctions are a bioelectric patterning element that sets up domains of isopotential cell fields during morphogenesis.
Serotonin signaling is used in information exchange between cells in processes such as L-R patterning and control of timing and cell movement during gastrulation. We have shown that serotonin is utilized by both chick and frog embryos, at very early stages, as a small molecule signal which is transported in a left-right gradient and regulates the development of laterality. Indeed, we now know that the early frog embryo is literally an electrophoresis chamber, which uses voltage potentials to generate consistently biased left-right gradients in serotonin in an epigenetic process not dependent on zygotic gene expression. We have modeled this process quantitatively, and characterized novel intracellular serotonin-binding proteins which directly activate asymmetric gene expression after their rightward movement, linking an early biophysical process to transcriptional regulation via chromatin modification pathways. Serotonin is also a key mediator for bioelectric control of neuronal outgrowth from transplants.
Electrical Fields Guiding 3D Shape of Cells and Organs
How does the cell know what size and shape it should be? Many cells alter their shape to provide different functions, like microglia. Even more complex is the question as to how organs, limbs, and creatures know what size and shape they should be when they are growing. How do the cells know how and where to form an organ? How does the creature know how to rebuild a severed limb to the correct size, shape, and orientation?
There are thousands of these same questions, including how astrocytes and neurons know the exact networks they should form. How immune cells know how to travel through very complex differing 3D environments? An equally difficult question has been addressed in previous posts as to how the
cell knows exactly what shape a coded sequence in a protein will take. Both cells and microbes alter the codes of their protein toxins that require extremely detailed and accurate shapes. In fact, modern science cannot calculate what shape a 400 amino acid coded sequence will be when it folds into a protein. It would take all the supercomputers together two thousand years to calculate the folding of one average protein. Yet, proteins assemble into the exact shape in a millisecond, helped by very complex chaperone molecules. Cells routinely edit their messenger RNA with alternative splicing to make a whole variety of shapes. How do they know how to do this?
Research shows that: If a cell of a two cell embryo is removed it still becomes the full creature.
Starved flatworms shrink keeping perfect proportions among the organs
Planaria can reproduce the entire body from a small piece
Amphibians can grow a perfectly proportioned limb
Many structures need nerve-mediated information in order to maintain shape and function
For all of these questions, it is hard to imagine the information of the 3D shape being inside one cell, or even a group of cells. It seems more likely that a field of information somehow guides these processes. Is this evidence for an electrical field of information forming 3D shapes of cells, organs and creatures, also, related to the influence of mind?
Electrical Gradients and Fields In and Around Cells

Previous posts have demonstrated the critical part that electrical synapses play in forming the structure of the neuronal network that uses chemical synapses. Also, a previous post showed that field potentials in and around brain cells, while poorly understood, are important for specific functions. Multiple posts have documented the elaborate communication that occurs between cells through signaling including neurotransmitters, brain factors, cytokines and hormones.
A significant amount of research shows the importance of various gradients in the space between cells for communication. This was noted in the posts on the movement of platelets that attract and signal to other immune cells. Another post showed the elaborate travels of leukocytes. A great deal is now known about morphogen gradients in the developing embryo—how cells know where to go and what types of cells to become.
Recent research shows that electric potentials in non brain cells are a signal for creating patterns during development and during re building of organs. This information of different field potentials surrounding individual cells can give information for the developing organ. Now, research from the laboratory of Dr. Michael Levin is demonstrating new ways that cells signal with electricity and the great importance of electrical properties for individual cells and tissues. He describes how electrical gradients and fields are critical in the 3D function and shape of cells and organs. Professor Levin notes that cancer cells use electric gradients for many purposes. In fact, aberrations in electrical processes can lead to cancer. Differences in electrical potential in cells are important in determining whether a cancer metastasizes and electrical states in one cell can have effects on distant cells, triggering metastasis. These electrical potentials in one cell can signal other cells that trigger genetic networks and epigenetic effects.
Electrical State of Cells
The electric state of a cell is determined by many complex factors. One is the opening and closing of ion channel proteins, which control the movement of charged particles (ions) across the cell membrane. Changes of the cells’ electrical potential via the activity of ion channels are shown to be able to suppress or trigger cancer. Another factor is electrical synapses that transfer electricity from one cell to another.
Different electrical properties of cells affect what type of cells are made from stem cells, how much reproduction goes on, where cells travel to and their shape and orientation. Electrical gradients and fields certainly affect travelling immune cells, stem cells, and all brain cells. They also exert profound effects on other somatic cell types during embryonic development and regenerative repair. Electrical properties are simultaneous with chemical metabolism using genetic network processing. They are critical for healing wounds and responding to infections. They determine the symmetry and shape of organs.
Differential ion channel activity in the body sets up specific patterns of voltage potential throughout tissues. During development, these electrical properties of cells determine what types of cells they become. For example, two specific groups of cells in the embryo have increased electrical potential and these become the two eyes. Research shows that by changing the electrical potential of cells at a critical time when eyes are being made, eyes can be induced to form elsewhere, effectively reprogramming other cell types into a complete visual organ.
While electrical activity can drive downstream genetic changes, just changing the electrical gradient can dramatically change the cell’s behavior. After an amputation, controlling the voltage potentials of wound cells causes the regrowth of a limb. The specification of the details appears to exist in the electrical field. At specific regions of the animal’s body, either a tail or limb grow due to the electrical signaling, which kickstarts a “build whatever goes here” program.
Electrical fields that are being discovered with multiple influences could contain this 3D information. Altering the electric field alters the information and the biological results.
Signaling with Chemicals and Electricity
Quite a number of previous posts reveal the elaborate signaling of microbes and immune cells such as T cells and platelets. The critical intestinal epithelial cells and skin cells are involved in very elaborate signaling between large numbers of microbes on one side and many immune cells on the others. Of course, brains are known to use elaborate signaling.
A previous post noted that cancer is like a microbe colony in that they work together for their ends. This view of cancer is that many cells in an environment trigger the actions of the community of cancer cells. Another view of cancer is as a defection of individual cells from the goals of the organism; this can occur by cells’ being isolated from, or mis-interpreting, the patterning cues that normally orchestrate cell behavior into complex anatomy. The new research by Levin adds a very large new dimension in the way electrical signals between cells might be critical in this cancer development.
A prominent aspect of multi cellular creatures is that they have organs of a particular size and shape. When regeneration occurs in reptiles the same exact shape is grown. Information for the cellular activity appears to exist in the space that will make up the specifically shaped organ. Levin notes that “cancer can be seen as an error of geometry, because tumor cells grow, migrate, and function without regard for the orderly structure within which they occur.”
This can occur through each cell having specific electrical gradients and properties that together form a large electric field of information. This field of information can show individual cells in the embryo how to behave. This is analogous to the fact that electrical flow between cells in the early embryo (see post on electrical synapses) forms the basic network that is then built into a formal structure with elaborate chemical synapses. This, also, occurs during rebuilding of tissue. Somehow, the information of the electrical flow through the electric gap junction synapses determines the future structure.
Electrical signals now appear to be critical in forming the shape of organs, the very function and identity of organs, and the creation of new limbs on animals that regenerate. In these animals, stem cell behavior is directed by currents created with potassium, sodium, chloride, and protons that affect the genetic networks of cells at a distance. Recent research shows that limbs can be influenced by proton and sodium alterations. New types of ion channels, pumps and electrical connections have now been found in a variety of different organs.
This is remarkable. If electric gap junction synapses determine cell shape, then macro change attributed commonly to macroevolution and structural biological novelties should also depend on these.
Voltage Gradients in Non Brain Cells
Cells not in the brain, also, use ion channels that create an electrical potential between the inside and outside of the cell. These do not send a signal down a long axon wire as neurons do, but the electrical potentials are critical for the cell’s behavior. In a previous post, calcium cyclic signaling was noted to be the crucial signal between plants and microbes when forming nitrogen factories.
Whole tissues and sheets of cells, also, have flows of electricity through electrical synapses called gap junctions (just like neurons). When the skin is broken the electrical gradients and fields are disrupted and these are a signal for immune cells to repair the tissue. An even larger field exists over an entire organ, and this field has crucial information regarding the shape and function of the organ.
It is very difficult to study, but even inside the cell, many membranes exist for the nucleus, the mitochondria and other organelles such as the critical information center of the endoplasmic reticulum. Electrical potentials and information fields exist in these, also.
Cancer and Electrical Gradients
Cancer cells’ behavior is very tied to electrical signals. They use them as clues to avenues of travel. Electrical fields determine shape changes in the cells. The electrical potential, also, affects what type of cell is produced from stem cells.
Cancer cells have altered electrical activity and use different ion channels and pumps. They have different transporter molecules that affect how the ions travel and, therefore, the electrical gradients. It now appears that these alterations in ion channels are signatures of different cancers and are critical to their formation. Many differences in the effects of cytokines and neurotransmitter signals, also, appear to be tied to these electrical differences. These ion channels influence all aspects of metastasis. In fact, oncogenes (special genes that cause a cell to become cancerous) are related to various different ion channels.
Research in cancer has been increasingly difficult because of the many different mutations that are found in types of cancers, and, more recently, in individuals. The studies have shown that there is a wide range of different mutations that don’t fall into any noticeable patterns.
Dr. Levin’s work points in a different direction entirely. He notes that the gradients and fields of electrical information may be a more important way to understand this process. It is the very space that the cells are living in that appears to be aberrant and causing the cancers. Electrical signaling is key for cells to properly interpret their environment, and when this process goes awry, the cells default to a cancer program.
Studies show that cancer cells can be differentiated from other cells by their electrical properties. The overall electrical status of the cancer cell is determined by the sum of all the ions and pumps. There are a vast amount of different possible genetic scenarios in this basic measurement of electrical status.
But, in fact, the simple electrical status appears to identify cancer cells. Dr. Levin likens this to “pressure” in physics – a group property that can be implemented with a wide variety of underlying molecular details (e.g., different ion channel genes). He notes that the statistical electrical study of cancer electric gradients might be the most information. He notes that the extremely complex analysis of all of the ion channels might not even reveal the critical information (this type of research is currently not even feasible).
Just analyzing messenger RNA and specific proteins made will not determine all the factors altering the global electrical fields because electrical state is a function of the 3D open/closed states of the channels, not merely their presence at the mRNA and protein levels.
The large electrical fields themselves appear to create the regulation that occurs. A study showed that by increasing the electric potential in cancer cells, metastatic activity can be suppressed, despite specific proteins being present that are known to be involved in metastatic activity.
Information Fields and Geometry
All of the signals that affect a cell, together, are called the morphogenic field. It includes all of the instructions that come from all secreted cellular signals, including cytokines and the effects of all the molecules in the extra cellular space (see post on Extracellular Space and Neuroplasticity). This includes many gradients that are used by immune cells to gather information and attract other cells. An unusual feature recently discovered as a cause of cancer is special proteins that produce cells in particular orientations—called planar cell polarity. When this polarity is altered it can cause cancer.
Electrical gradients are now shown to be critical as well. When the usual gradient or field information is somehow altered it can cause cancer. Each embryo has its own innate field. It has been observed that cells in the midst of a live organ are not as susceptible to the same chemical influences to become cancerous. They are much more able to become cancer when isolated from the influences of other cells. When the region’s electrical properties are different, they are less likely to become cancerous.
Signals from neurons are, also, a factor in causing cancer. When nerves are cut, then cancers occur more frequently. These neuron signals appear to be contributing to the field information in local regions.
A striking example of the field is when shunts are needed between the abdomen and the blood. These shunts send large amounts of cancer cells to many regions, but they do not become metastatic in some areas, but do in others. Another is when the shape of a region is altered by surgery it is more likely to become cancerous, possibly by altering the field.
Cancer cells turn away from the community aspect of the entire organism and become involved in individual behavior and their own new community (see post on Cancer – The Emperor of Cells.) The cancer cells either aren’t given appropriate information in the field about the creature, can’t extract the information or don’t care anymore. Cancers form their own organs and fields. The cancer cells, also, form their own information fields and communicate and cooperate using them. They reproduce and evolve to be stronger as a community. They signal each other to fight microbes and immune attacks. Cancer cells eliminate communication to the body and tissues of the body. But, they maintain the same junctions and communications among this smaller community.
Where is the Information
The study of electric fields brings up the problem of orders of magnitude of effect and the question of emergent properties. While at the level of genetic network activity, each cell in the field has definite behaviors—they are all influenced by the overall information fields. All levels are at play—the genetic, epigenetic, individual cells and networks of cells.
Having information in morphogenic fields including electrical gradients is similar to the notion that mental events occur in neuronal electric networks. Dr. Levin speculates that the same way that information is contained in neural networks, information could be maintained in electrical fields of cells, organs and creatures.
All of this, raises the question, of course, about its relation to cellular intelligence in particular and mind in nature in general.
Which Comes First – Electric Fields or Biochemistry
The genetic networks are very critical in electric signaling because cells use protein ion channels imbedded in the membrane. These proteins affect distant actions and are very involved in creating the overall gradients.
In one of many circular genetic processes, the actions of the genetic machinery are controlled by electrical gradients and the elaborate signaling of the genetic networks create the electrical gradient.
Nices Catch22 situation.
Patterning occurs by a continuous interplay of genetics and physics.
This conundrum indicates that there is a more global regulation of the entire process. In fact, research shows that the electrical influences cannot be reduced to simple chemistry inside of a cell. Electrical gradients of individual cells and many cells contribute to the creation of the 3D shapes and functions of organs and creatures. Examples of this include the creation of the embryonic eye, the head, and the regeneration of limbs exactly to size. For movement of cells, when electrical and chemical gradients are in competition, the electrical gradient wins.
The example of the eye is instructive. Dr. Levin notes that it shows the electrical information is more crucial. It is the electrical properties of the regions forming the two eyes that determine where the eyes will go. When this electrical property is altered, the eye forms in different places. This effect cannot be replicated outside of the head by any of the known “master” eye genes.
What Signals Could Describe 3 Dimensions?
How does a cell first create a shape of an organ in the fetus and then maintain it throughout life, with many new cells being generated? Signals can let a cell know about position, size and polarity and orientation. It is either cell-to-cell communication, which is basically the current research goal, or a pre formed field or organizing template. The 3D process in the fetus is called morphogenesis. Each cell in the embryo has to know its position relative to others and differentiates into the appropriate type of cell for that position.
Field structures in physics are similar to the concept of morphogenetic field. A field includes information at each location. It includes regulation at a distance. The gradient is a concept already used in studies of development. Current research shows that electrical potentials and gradients in the space between cells are an important source of information for cells, and control the movement of chemical signals such as serotonin and butyrate.
Electrical Fields Guiding 3D Shape of Cells and Organs
While both chemical and electrical factors are clearly relevant to determining the three dimensional function and shapes of cells, organs and creatures, it appears that an information field of some kind would be necessary for the level of information needed. Recent research shows the importance of the electrical potentials, gradients and fields.
Among theories of what mind could be (see post), information fields have been proposed. Perhaps this current research furthers such a view.
1) http://reasonandscience.heavenforum.org/t1826-how-does-evolution-supposedly-work
2) http://reasonandscience.heavenforum.org/t2279-what-prevents-the-transition-from-micro-to-macro-evolution
3) http://jonlieffmd.com/blog/electrical-fields-guiding-3d-shape-of-cells-and-organs
4) http://www.drmichaellevin.org/research.html
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3413735/
Last edited by Otangelo on Mon Aug 01, 2022 4:51 pm; edited 15 times in total