Primary Cilium a Cell’s Antenna or Its Brain
https://reasonandscience.catsboard.com/t2089-primary-cilium-a-cells-antenna-or-its-brain
The argument from wound healing cilium
1. The cilium that looks like an antenna on most human cells, orients cells to move in the right direction at the speed needed to heal wounds, and so acts like a Global Positioning System (GPS) that helps ships navigate to their destinations.
2. “The really important discovery is that the primary cilium detects signals, which tell the cells to engage their compass reading and move in the right direction to close the wound.”
3. “Protruding through the cell membrane, primary cilia occur on almost every non-dividing cell in the body.”
4. “Once written off as a vestigial organelle discarded in the evolutionary dust, primary cilia in the last decade have risen to prominence as a vital cellular sensor at the root of a wide range of health disorders, from polycystic kidney disease to cancer to left-right anatomical abnormalities.”
5. The unavoidable importance of the primeval cilium for the survival of the cell and its wonderful design proves the existence of the primeval designer God.
6. God necessarily exists.
The flagellum is a prima facie example of intelligent design, which has become well known since Behe's book Darwins Blackbox. In his following book, The edge of evolution, Behe describes in detail the Cilia, the non-motile sister of the flagellum. It is less known. What science has discovered in recent years, is a serious challenge for proponents of naturalistic origins. Jon Lieff describes the amazing capabilities of the cilia in his article: 1

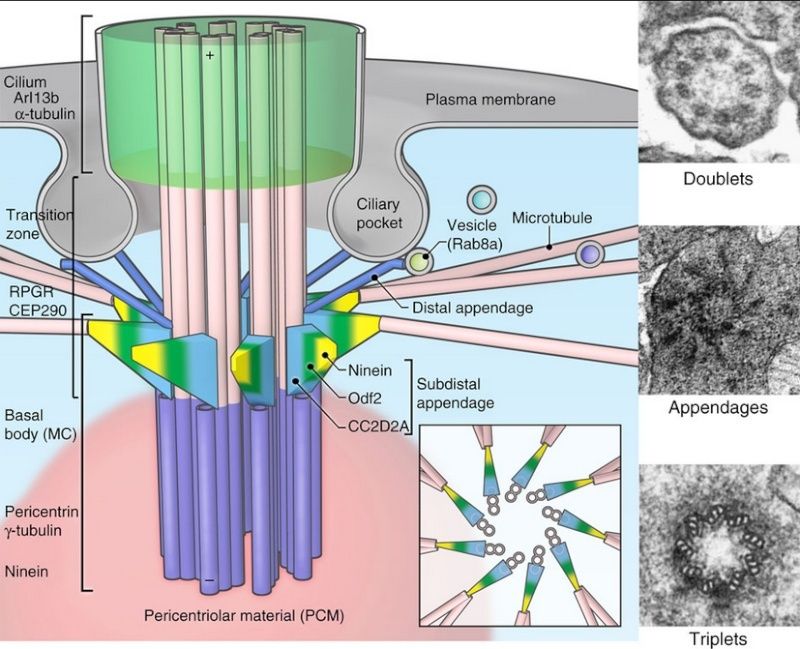
Given the sophistication of the cilium’s function, it is not surprising the cilium is one of the most structurally complex organelles in the cell. 2
Almost every human cell has a little-known structure called the primary cilium. It is similar to the well known motile cilia, but without special structures for movement. This solitary, unmoving structure, most often sticking out of cells, was considered a vestigial organ. While first noted in 1987, only recently has the primary cilium been proven to have very important sensory functions—an antenna receiving information about events external to the cell. More recently it has been found to be critical for cellular communication and signaling in fetal development of all cells, but particularly brain cells. Specialized versions of the primary cilium appear in key places for sensation, such as receptors for hearing, sight and smell. Recently, a close link was found to the all-important centriole spindle that directs all cell division. Equally unusual is the discovery of a very complex theinheritanunique transport system within the primary cilium, whose defects now account for many important diseases. Is the primary cilium a cell’s antenna or its brain?
Ninety-nine percent of mammal cells have a protrusion of the outer membrane, called the primary cilium. This includes all neurons and glial cells. Unlike other cilia, it is not an organ for movement. Abnormalities in the primary cilium cause retinitis pigmentosa, hydrocephalus, polycystic kidney disease, malformations of the skeleton, defects of the neural tube and some obesity.

Cilia and Flagella
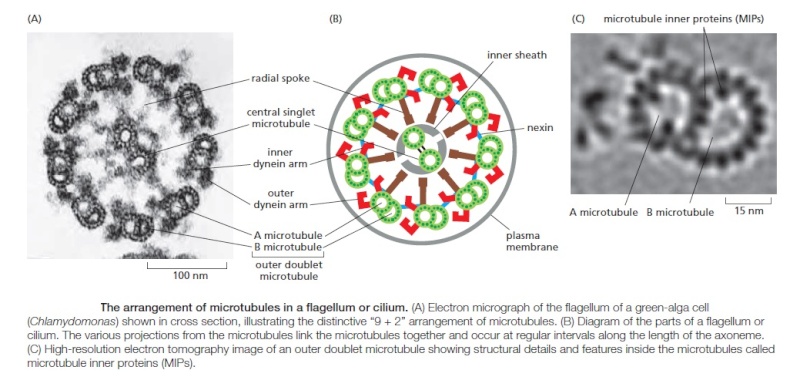
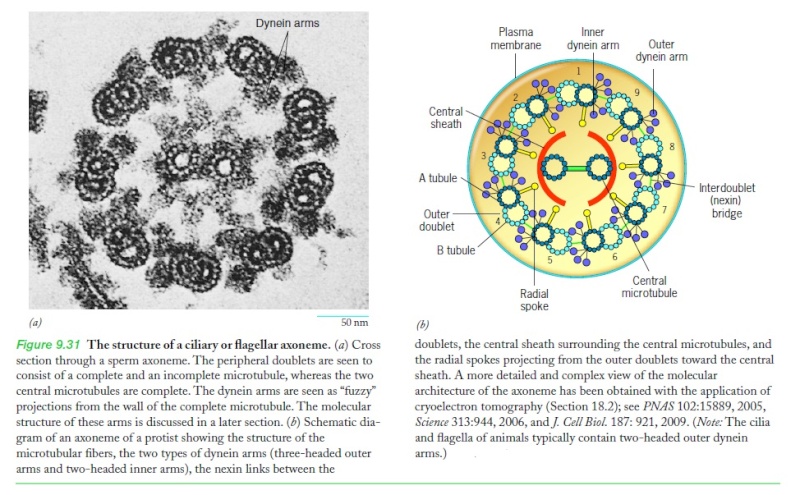
Cellular protrusions or tails have been called flagella and cilia. Flagella are long and cilia are short. Flagella and mobile cilia have the same structure designed for movement with cross-connections between 9 primary microtubules. The primary cilium does not have the Sperm connections for movement. When flagella are present in a cell, there is a small number—less than 5. There can be over a hundred cilia in rows.
Sperm cells are mostly a tail that moves; it is a very long flagellum. Some cells have rows of many cilia that often function in a synchronized movement. Cells that line the surface of the lungs and respiratory passages have 200 cilia all moving and beating using a complex swimming motion with a stroke for power and then one for recovery. These cilia move water and mucous along the tubes. In the ear hair cells have Axeneme detail many cilia that pick up pressure and sound. Cilia propel fluid in the CSF to circulate in the ventricles through pores to spinal cord, brain stem, cerebellum and cortex.
Bacterial cilia and flagella have many different structures compared with the one structure in all eukaryote cell. Each flagella bacteria species have many different structures, with widely varying proteins.
All bacterial flagella work differently than the tails of animal and human cells. Bacteria have rotating flagella like propellers—turning three to fifteen times per second. With this propeller machinery, they move very rapidly—up to 50 cell lengths per second.
Swimming algae synchronize their two large flagella making a movement like a breaststroke. In order to maintain synchronization, the algae rocks, which speeds up or slows down the movement. While it appears to be simple, this movement involves ten thousand complex molecular machines working together.

The Primary Cilium – PI
For a hundred years it was thought that the beating movement was the main function of cells’ tails. Recently they are noted to be antenna with sensory machinery. With no center microtubules, the primary cilium is a unique environment 1/10,000th the size of the cell. Because it is a circumscribed small area with a very specific diameter, unusual neuron cilium autismproteins can anchor there and perform unique functions. There are many different sensory receptors in the membrane responding to the environment, tracking mechanical and chemical forces and sending signals to the organism. Some properties of the membrane are unique in the primary cilium. Because it is separate from the rest of the cell, special proteins can accumulate 100 times more than other places and make the signaling much more efficient.
The single, non-beating primary cilia have many different receptors and very important functions. In the kidneys the primary cilium responds to the flow of the liquid through tubules. Bending from the pressure triggers calcium signaling and is part of critical kidney regulation. smell receptor primary cilium are now known to sense different chemicals, concentrations of ions, temperature and gravity. In the nose the olfactory receptors are in primary cilium. In the eye, the light-sensing receptor is an outpouching at the tip of the primary cilium. The PI senses light wavelengths in eye cells, pressure in cartilage, and blood flow in heart cells.In the ear, cilia sense vibrations. Although it was thought that mobile cilia were present in many cells, in fact, more cells have non-mobile primary cilium.
An important recent study shows that blocking receptors on the primary cilium in mice causes memory loss. A protein (PQBP1) was identified that if absent eliminates the PI in neurons causing problems with memory and learning. PQBP1 is usually found only in the cell’s nucleus, but in the neuron, it is in the base of the primary cilium. PQBP1 appears to bind to another protein that normally would suppress the cilium. It stops the suppression, allowing the creation of the PI in the neuron. PI has also been shown to be critical for fetal brain development. Moving cilia are critical to migration of neurons and glia in developing brain. But, the non-moving PI is involved in communication with other cells during these migrations. In the hippocampus, where new brain cells originate in adults (neurogenesis), PIs are necessary for stem cells making new neurons. When receptors in the PI respond to the external environment of the cell, receptors activate cascades that communicate to the cell nucleus using special transport motors. Without the PI in neurons during development, several brain diseases occur.
Transport System
The primary cilium is a hair-like structure that protrudes from the cell surface. Microtubules form the core structure of the cilium, the axoneme. Protein cargo is transported up and down the cilium via anterograde and retrograde IFT mediated by kinesin and dynein motor proteins, respectively, which travel along the axoneme.
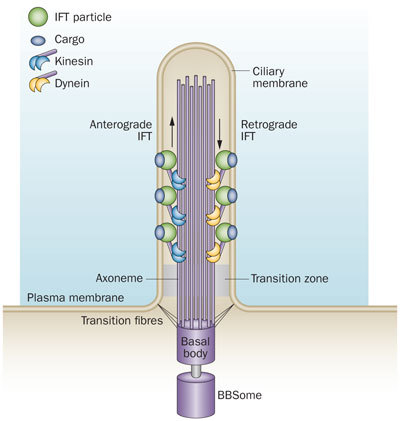
the intra-flagella transport system IFT in the PI was observed and connected to polycystic kidney disease. It was observed that the microtubule structure, called axoneme, grows from the base of the primary cilium, called the basal body. When the cilium is being built, vesicles transport protein pieces to the growing cilium from the base.
Recently, the complexity of the transport system has been described. A special protein attaches and drags proteins through the sea of phospholipids in the membrane, and pulls them into the primary cilium . In this way the primary cilium becomes the communication hub of the cell. Already a number of critical signaling cascades have been shown to live in the primary cilium. This includes the critical hedgehog and retinal signaling pathways.

The transport system uses motors that travel along microtubules to get the important material to the tip of the primary cilium from the base. Special motors are built at the base of the primary cilium and they pull many different types of material into the primary cilium—receptor proteins and building blocks for microtubules.Once at the tip of the PI, the motors deposit the cargo. At the tip Kinesian color the motor is altered and becomes a different machine to bring signaling material down the PI to the base. At the based messages are created and sent to the nucleus.
Once at the base of the primary cilium, the motor rearranges itself and becomes the train that drives cargo up into the primary cilium. The train that pulls this material to the tip of the primary cilium is made of at least four motors, one type active at a time. These motors are not just motors; they also interact with the membrane to regulate other functions including sensing extracellular situations and influencing decisions during fetal development. These motors are also able to connect through the membrane to objects outside of the cell to stimulate different types of cell movement. In this situation, the motor is anchored to a particular spot, but the entire cell moves when the motor is turned on.
This very complex motor system is critical for the elaborate function of the primary cilium by transporting all receptors and signaling materials that are used for the antenna function.
Defects in these motors in the eye can cause blindness. In the cells of the eye, the tip of the PI is very large bulb and houses the sensors that respond to light but still has a narrow area connecting with the large cell body. All of the light signals have to go through this narrow tube and a defect leads to blindness. Proteins used for sensing light are fragile and very active and many are imperfect. The cell is very dependent on the transport system of the PI to continually restock the proteins. Retinitis pigmentosa is one of the many diseases (ciliopathies) related to defects in this transport system.
T Cell Cilium
Until recently, it was not obvious that lymphocytes have PIs. Now, it has been shown that when T cells form critical synapses with other cells—other T cells for activation, dendritic cells to receive antigens, other immune cells for regulation, and targeted cells for destruction—it is actually a PI variant in the T cell.
The base of the cilium consists of a complex microtubule structure called the microtubule-organizing center, or MTOC (also called a centrosome). This is where microtubules are built. This MTOC becomes the focus of the synapses T cells form with other cells for these critical functions. The same PI transport system is used to transport vesicles during these close encounters with other cells. Vesicles are transported by the cilium transport system to the membrane to kill the targeted cell. The transport system brings special receptors that are recycled during T cell activation. T cells, like PIs, also, use the hedgehog signaling process during activation.
Autophagy and PI
Another critical function in all cells is the pathway of growth and the opposite, cellular self-destruction. The autophagy process is a critical quality control for communities of cells, such as organs. Neurodegenerative disease and cancer are highly connected to cascades that lead to runaway growth at one end and cell self-destruction on the other.
Signaling from the primary cilium, such as using the Hedgehog pathway, appears to induce autophagy. Autophagy and PIs both utilize the same set of complex molecules. It has been found recently that the PI is necessary for stimulation of autophagy. The autophagy machinery is at the base of the PI. The PI regulates autophagy and the autophagy proteins direct the receptors in the PI. All critical sensory and communicating functions are connected with both the PI and autophagy mechanism.
Is the Primary Cilium a Cell’s Antenna or Its Brain?
the primary cilium is the brain of a cell, then it raises the question as to how the critical microtubule structures relate to this. The Penrose-Hammerof theory of the biological basis of mind is based on quantum properties of microtubules. In this theory, unique quantum computing processes occur in the exact unusual structure of microtubules. Currently, this theory is unable to be proven but does provide a speculation as to how the all-important mitotic spindle and the primary cilium might serve as a brain for the cell.
Previous posts have described intelligence in microbes. Microbes swim, communicate, find food, and perform group actions without synapses. All of these sensing and movement behaviors involve microtubules. In the human neuronal synapse, neuroplasticity ( refers to changes in neural pathways and synapses due to changes in behavior, environment, neural processes, thinking, and emotions - as well as to changes resulting from bodily injury ) depends upon the constantly changing microtubule structures. Just like the primary cilium and the mitotic spindle, purposeful complex behavior at many levels appears to involve the microtubules. The extremely complex mitotic spindle in cell division includes a small sac of the PI material. Is this sac passing on specific information encoded in the microtubule structure for the next generation?
With the primary cilium at the center of cellular sensation, communication, movement, cell division, autophagy and decision making, it is clearly a hub of purposeful behavior in all cells including the neuron. Is it possible that the primary cilium is the brain of the cell?
1) http://jonlieffmd.com/blog/is-the-primary-cilium-a-cells-antenna-or-its-brain#sthash.VkxokOmV.dpuf
2) http://jcs.biologists.org/content/123/4/499
https://reasonandscience.catsboard.com/t2089-primary-cilium-a-cells-antenna-or-its-brain
The argument from wound healing cilium
1. The cilium that looks like an antenna on most human cells, orients cells to move in the right direction at the speed needed to heal wounds, and so acts like a Global Positioning System (GPS) that helps ships navigate to their destinations.
2. “The really important discovery is that the primary cilium detects signals, which tell the cells to engage their compass reading and move in the right direction to close the wound.”
3. “Protruding through the cell membrane, primary cilia occur on almost every non-dividing cell in the body.”
4. “Once written off as a vestigial organelle discarded in the evolutionary dust, primary cilia in the last decade have risen to prominence as a vital cellular sensor at the root of a wide range of health disorders, from polycystic kidney disease to cancer to left-right anatomical abnormalities.”
5. The unavoidable importance of the primeval cilium for the survival of the cell and its wonderful design proves the existence of the primeval designer God.
6. God necessarily exists.
The flagellum is a prima facie example of intelligent design, which has become well known since Behe's book Darwins Blackbox. In his following book, The edge of evolution, Behe describes in detail the Cilia, the non-motile sister of the flagellum. It is less known. What science has discovered in recent years, is a serious challenge for proponents of naturalistic origins. Jon Lieff describes the amazing capabilities of the cilia in his article: 1

Given the sophistication of the cilium’s function, it is not surprising the cilium is one of the most structurally complex organelles in the cell. 2
Almost every human cell has a little-known structure called the primary cilium. It is similar to the well known motile cilia, but without special structures for movement. This solitary, unmoving structure, most often sticking out of cells, was considered a vestigial organ. While first noted in 1987, only recently has the primary cilium been proven to have very important sensory functions—an antenna receiving information about events external to the cell. More recently it has been found to be critical for cellular communication and signaling in fetal development of all cells, but particularly brain cells. Specialized versions of the primary cilium appear in key places for sensation, such as receptors for hearing, sight and smell. Recently, a close link was found to the all-important centriole spindle that directs all cell division. Equally unusual is the discovery of a very complex theinheritanunique transport system within the primary cilium, whose defects now account for many important diseases. Is the primary cilium a cell’s antenna or its brain?
Ninety-nine percent of mammal cells have a protrusion of the outer membrane, called the primary cilium. This includes all neurons and glial cells. Unlike other cilia, it is not an organ for movement. Abnormalities in the primary cilium cause retinitis pigmentosa, hydrocephalus, polycystic kidney disease, malformations of the skeleton, defects of the neural tube and some obesity.

Cilia and Flagella
Cellular protrusions or tails have been called flagella and cilia. Flagella are long and cilia are short. Flagella and mobile cilia have the same structure designed for movement with cross-connections between 9 primary microtubules. The primary cilium does not have the Sperm connections for movement. When flagella are present in a cell, there is a small number—less than 5. There can be over a hundred cilia in rows.
Sperm cells are mostly a tail that moves; it is a very long flagellum. Some cells have rows of many cilia that often function in a synchronized movement. Cells that line the surface of the lungs and respiratory passages have 200 cilia all moving and beating using a complex swimming motion with a stroke for power and then one for recovery. These cilia move water and mucous along the tubes. In the ear hair cells have Axeneme detail many cilia that pick up pressure and sound. Cilia propel fluid in the CSF to circulate in the ventricles through pores to spinal cord, brain stem, cerebellum and cortex.
Bacterial cilia and flagella have many different structures compared with the one structure in all eukaryote cell. Each flagella bacteria species have many different structures, with widely varying proteins.
All bacterial flagella work differently than the tails of animal and human cells. Bacteria have rotating flagella like propellers—turning three to fifteen times per second. With this propeller machinery, they move very rapidly—up to 50 cell lengths per second.
Swimming algae synchronize their two large flagella making a movement like a breaststroke. In order to maintain synchronization, the algae rocks, which speeds up or slows down the movement. While it appears to be simple, this movement involves ten thousand complex molecular machines working together.

The Primary Cilium – PI
For a hundred years it was thought that the beating movement was the main function of cells’ tails. Recently they are noted to be antenna with sensory machinery. With no center microtubules, the primary cilium is a unique environment 1/10,000th the size of the cell. Because it is a circumscribed small area with a very specific diameter, unusual neuron cilium autismproteins can anchor there and perform unique functions. There are many different sensory receptors in the membrane responding to the environment, tracking mechanical and chemical forces and sending signals to the organism. Some properties of the membrane are unique in the primary cilium. Because it is separate from the rest of the cell, special proteins can accumulate 100 times more than other places and make the signaling much more efficient.
The single, non-beating primary cilia have many different receptors and very important functions. In the kidneys the primary cilium responds to the flow of the liquid through tubules. Bending from the pressure triggers calcium signaling and is part of critical kidney regulation. smell receptor primary cilium are now known to sense different chemicals, concentrations of ions, temperature and gravity. In the nose the olfactory receptors are in primary cilium. In the eye, the light-sensing receptor is an outpouching at the tip of the primary cilium. The PI senses light wavelengths in eye cells, pressure in cartilage, and blood flow in heart cells.In the ear, cilia sense vibrations. Although it was thought that mobile cilia were present in many cells, in fact, more cells have non-mobile primary cilium.
An important recent study shows that blocking receptors on the primary cilium in mice causes memory loss. A protein (PQBP1) was identified that if absent eliminates the PI in neurons causing problems with memory and learning. PQBP1 is usually found only in the cell’s nucleus, but in the neuron, it is in the base of the primary cilium. PQBP1 appears to bind to another protein that normally would suppress the cilium. It stops the suppression, allowing the creation of the PI in the neuron. PI has also been shown to be critical for fetal brain development. Moving cilia are critical to migration of neurons and glia in developing brain. But, the non-moving PI is involved in communication with other cells during these migrations. In the hippocampus, where new brain cells originate in adults (neurogenesis), PIs are necessary for stem cells making new neurons. When receptors in the PI respond to the external environment of the cell, receptors activate cascades that communicate to the cell nucleus using special transport motors. Without the PI in neurons during development, several brain diseases occur.
Transport System
The primary cilium is a hair-like structure that protrudes from the cell surface. Microtubules form the core structure of the cilium, the axoneme. Protein cargo is transported up and down the cilium via anterograde and retrograde IFT mediated by kinesin and dynein motor proteins, respectively, which travel along the axoneme.

the intra-flagella transport system IFT in the PI was observed and connected to polycystic kidney disease. It was observed that the microtubule structure, called axoneme, grows from the base of the primary cilium, called the basal body. When the cilium is being built, vesicles transport protein pieces to the growing cilium from the base.
Recently, the complexity of the transport system has been described. A special protein attaches and drags proteins through the sea of phospholipids in the membrane, and pulls them into the primary cilium . In this way the primary cilium becomes the communication hub of the cell. Already a number of critical signaling cascades have been shown to live in the primary cilium. This includes the critical hedgehog and retinal signaling pathways.

The transport system uses motors that travel along microtubules to get the important material to the tip of the primary cilium from the base. Special motors are built at the base of the primary cilium and they pull many different types of material into the primary cilium—receptor proteins and building blocks for microtubules.Once at the tip of the PI, the motors deposit the cargo. At the tip Kinesian color the motor is altered and becomes a different machine to bring signaling material down the PI to the base. At the based messages are created and sent to the nucleus.
Once at the base of the primary cilium, the motor rearranges itself and becomes the train that drives cargo up into the primary cilium. The train that pulls this material to the tip of the primary cilium is made of at least four motors, one type active at a time. These motors are not just motors; they also interact with the membrane to regulate other functions including sensing extracellular situations and influencing decisions during fetal development. These motors are also able to connect through the membrane to objects outside of the cell to stimulate different types of cell movement. In this situation, the motor is anchored to a particular spot, but the entire cell moves when the motor is turned on.
This very complex motor system is critical for the elaborate function of the primary cilium by transporting all receptors and signaling materials that are used for the antenna function.
Defects in these motors in the eye can cause blindness. In the cells of the eye, the tip of the PI is very large bulb and houses the sensors that respond to light but still has a narrow area connecting with the large cell body. All of the light signals have to go through this narrow tube and a defect leads to blindness. Proteins used for sensing light are fragile and very active and many are imperfect. The cell is very dependent on the transport system of the PI to continually restock the proteins. Retinitis pigmentosa is one of the many diseases (ciliopathies) related to defects in this transport system.
T Cell Cilium
Until recently, it was not obvious that lymphocytes have PIs. Now, it has been shown that when T cells form critical synapses with other cells—other T cells for activation, dendritic cells to receive antigens, other immune cells for regulation, and targeted cells for destruction—it is actually a PI variant in the T cell.
The base of the cilium consists of a complex microtubule structure called the microtubule-organizing center, or MTOC (also called a centrosome). This is where microtubules are built. This MTOC becomes the focus of the synapses T cells form with other cells for these critical functions. The same PI transport system is used to transport vesicles during these close encounters with other cells. Vesicles are transported by the cilium transport system to the membrane to kill the targeted cell. The transport system brings special receptors that are recycled during T cell activation. T cells, like PIs, also, use the hedgehog signaling process during activation.
Autophagy and PI
Another critical function in all cells is the pathway of growth and the opposite, cellular self-destruction. The autophagy process is a critical quality control for communities of cells, such as organs. Neurodegenerative disease and cancer are highly connected to cascades that lead to runaway growth at one end and cell self-destruction on the other.
Signaling from the primary cilium, such as using the Hedgehog pathway, appears to induce autophagy. Autophagy and PIs both utilize the same set of complex molecules. It has been found recently that the PI is necessary for stimulation of autophagy. The autophagy machinery is at the base of the PI. The PI regulates autophagy and the autophagy proteins direct the receptors in the PI. All critical sensory and communicating functions are connected with both the PI and autophagy mechanism.
Is the Primary Cilium a Cell’s Antenna or Its Brain?
the primary cilium is the brain of a cell, then it raises the question as to how the critical microtubule structures relate to this. The Penrose-Hammerof theory of the biological basis of mind is based on quantum properties of microtubules. In this theory, unique quantum computing processes occur in the exact unusual structure of microtubules. Currently, this theory is unable to be proven but does provide a speculation as to how the all-important mitotic spindle and the primary cilium might serve as a brain for the cell.
Previous posts have described intelligence in microbes. Microbes swim, communicate, find food, and perform group actions without synapses. All of these sensing and movement behaviors involve microtubules. In the human neuronal synapse, neuroplasticity ( refers to changes in neural pathways and synapses due to changes in behavior, environment, neural processes, thinking, and emotions - as well as to changes resulting from bodily injury ) depends upon the constantly changing microtubule structures. Just like the primary cilium and the mitotic spindle, purposeful complex behavior at many levels appears to involve the microtubules. The extremely complex mitotic spindle in cell division includes a small sac of the PI material. Is this sac passing on specific information encoded in the microtubule structure for the next generation?
With the primary cilium at the center of cellular sensation, communication, movement, cell division, autophagy and decision making, it is clearly a hub of purposeful behavior in all cells including the neuron. Is it possible that the primary cilium is the brain of the cell?
1) http://jonlieffmd.com/blog/is-the-primary-cilium-a-cells-antenna-or-its-brain#sthash.VkxokOmV.dpuf
2) http://jcs.biologists.org/content/123/4/499
Last edited by Admin on Sat May 09, 2020 5:12 pm; edited 13 times in total