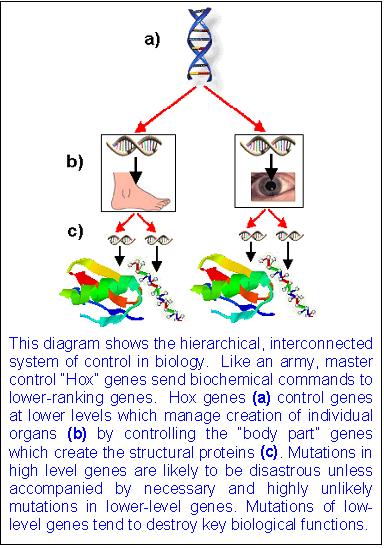
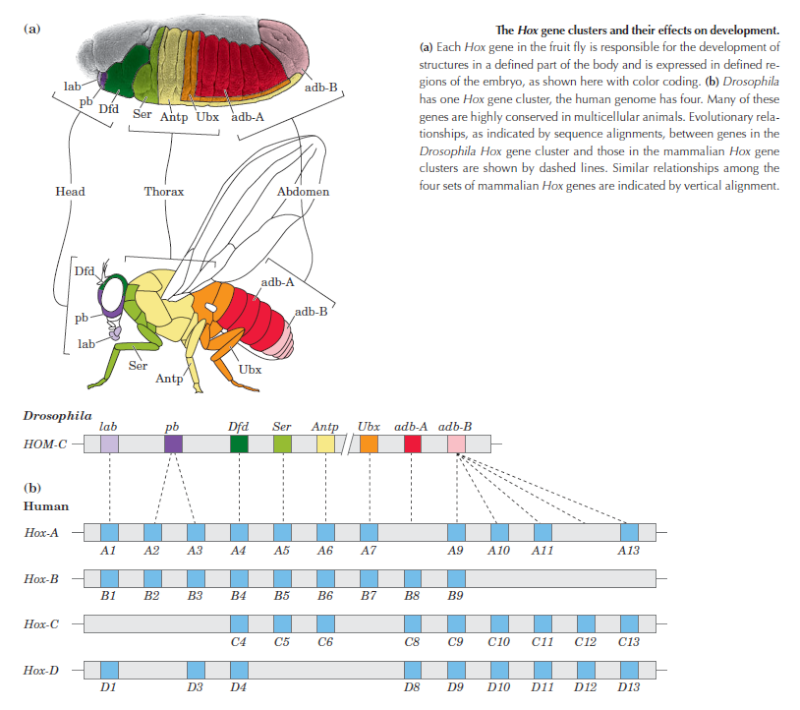
Hox Genes
http://www.researchgate.net/post/Why_do_researchers_continue_to_mislead_with_the_notion_that_Hox_genes_determine_the_body_plan
Genes code for proteins and RNAs. They don't code for brains, limbs or body plans. Yet many scientists insist that Hox genes, and other transcription factors, are responsible for "laying out the flooor plan" of the organism when all they are observed to do is activate genes in already established segments of the developing embryo. We have known this for 20 years. So why are the 39 Hox genes still portrayed as determining the geometry of anatomy when all they do is bind to DNA and RNA polymerase to affect gene expression in ontogeny?
Yes, mutations in hox genes do produce aberrant body plans..like legs where there should be antennae in fruit flies. But it is a mistake to assume that this is evidence that hox genes causally lay out the body plan, just like it would be a mistake to assume that a fault in a TV, causing a disruption in the reception of the signal, shows that the TV box produces the signal itself. Linear DNA cannot produce 3D form.
I am questioning th role of Hox genes in particular, as they are the most widely touted, but I am more generallty questioning the role of transcription factors and signalling proteins as a whole since their function is only to affect the production of other proteins, not to instruct cells how to form specific 3D structures like hearts and lungs. Yes, proteins are three dimensional entities, but there is no proof that their encoded amino acid sequence alone determines their final conformation - but that is not really an issue for developmental biology. My main argument is that the body plan of the organism is not encoded in DNA, or represented in the membrane/cytoskeletal patterns of the egg. Think about it: the human brain has 10^15 synaptic connections, which is 300,000 times more than the number of base pairs in the entire genome! There is a clear difference between genomic complexity and the complexity of the organism.
Darwins doubt, pg.239
WHAT ABOUT HOX GENES?
Hox (or homeotic) genes regulate the expression of other protein-coding genes during the process of animal development. Some biologists have likened them to the conductor of an orchestra who plays the role of coordinating the contributions of the players. And because Hox genes affect so many other genes, many evo-devo advocates think that mutations in these genes can generate large- scale changes in form.
But can mutations in Hox genes transform one form of animal life—one body plan—into another? There are several reasons to doubt that they can.
First, precisely because Hox genes coordinate the expression of so many other different genes, experimentally generated mutations in Hox genes have proven harmful. in fruit flies "most mutations in homeotic [Hox] genes cause fatal birth defects." In other cases, the resulting Hox mutant phenotype, while viable in the short term, is nonetheless markedly less fit than the wild type. For example, by mutating a Hox gene in a fruit fly, biologists have produced the dramatic Antennapedia mutant, a hapless fly with legs growing out of its head where the antennae should be
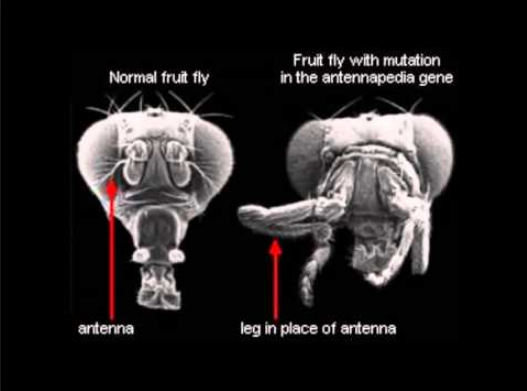
Other Hox mutations have produced fruit flies in which the balancers (tiny structures behind wings that stabilize the insect in flight, called "halteres") are transformed into an extra pair of wings. Such mutations alter the structure of the animal, but not in a beneficial or permanently heritable way. The Antennapedia mutant cannot survive in the wild; it has difficulty reproducing, and its offspring die easily. Similarly, fruit-fly mutants sporting an extra set of wings lack the musculature to make use of them and, absent their balancers, cannot fly. As Hungarian evolutionary biologist Eörs Szathmáry notes with cautious understatement in the journal Nature, "macromutations of this sort [i.e., in Hox genes] are probably frequently maladaptive."
Second, Hox genes in all animal forms are expressed after the beginning of animal development, and well after the body plan has begun to be established. In fruit flies, by the time that Hox genes are expressed, roughly 6,000 cells have already formed, and the basic geometry of the fly—its anterior, posterior, dorsal, and ventral axes—is already well established. So Hox genes don't determine body-plan formation. Eric Davidson and Douglas Erwin have pointed out that Hox gene expression, although necessary for correct regional or local differentiation within a body plan, occurs much later during embryogenesis than global body-plan specification itself, which is regulated by entirely different genes. Thus, the primary origin of animal body plans in the Cambrian explosion is not merely a question of Hox gene action, but of the appearance of much deeper control elements—
Davidson's "developmental gene regulatory networks" (dGRNs).Davidson argues that it is extremely difficult to alter dGRNs without damaging their ability to regulate animal development. Third, Hox genes only provide information for building proteins that function as switches that turn other genes on and off. The genes that they regulate contain information for building proteins that form the parts of other structures and organs. The Hox genes themselves, however, do not contain information for building these structural parts. In other words, mutations in Hox genes do not have all the genetic information necessary to generate new tissues, organs, or body plans.
Nevertheless, Schwartz argues that biologists can explain complex structures such as the eye just by invoking Hox mutations alone. He asserts that "[t]here are homeobox genes for eye formation and that when one of them, the Rx gene in particular, is activated in the right place and at the right time, an individual has an eye." He also thinks that mutations in Hox genes help arrange organs to form body plans. In a review of Schwartz's book, Eörs Szathmáry finds Schwartz's reasoning deficient. He too notes that Hox genes don't code for the proteins out of which body parts are made. It follows, he insists, that mutations in Hox genes cannot by themselves build new body parts or body plans. As he explains, "Schwartz ignores the fact that homeobox genes are selector genes. They can do nothing if the genes regulated by them are not there." Though Schwartz says he has "marveled" at "the importance of homeobox genes in helping us to understand the basics of evolutionary change," kind of (nonhomologous) leg. Another homologue of the Distal-less gene in echinoderms regulates the development of tube feet and spines—anatomical features classically thought not to be homologous to arthropod limbs, nor to limbs of tetrapods.34 In each case, the Distal-less homologues play different roles determined by the higher-level organismal context. And since mutations in Hox genes do not alter higher-level epigenetic contexts,35 they cannot explain the origin of the novel epigenetic information and structure that establishes the context and that is necessary to building a new animal body plan.
Szathmáry doubts that mutations in these genes have much creative power. After asking whether Schwartz succeeds in explaining the origin of new forms of life by appealing to mutations in Hox genes, Szathmáry concludes, "I'm afraid that, in general, he does not."Nor, of course, do Hox genes possess the epigenetic information necessary for body-plan formation. Indeed, even in the best of cases mutations in Hox genes still only alter genes. Mutations in Hox genes can only generate new genetic information in DNA. They do not, and cannot, generate epigenetic information. Instead, epigenetic information and structures actually determine the function of many Hox genes, and not the reverse. This can be seen when the same Hox gene (as determined by nucleotide sequence homology) regulates the development of different anatomical features found in different phyla. For instance, in arthropods the Hox gene Distal-less is required for the normal development of jointed arthropod legs. But in vertebrates a homologous gene (e.g., the Dlx gene in mice) builds a different
http://www.researchgate.net/post/Why_do_researchers_continue_to_mislead_with_the_notion_that_Hox_genes_determine_the_body_plan
Genes code for proteins and RNAs. They don't code for brains, limbs or body plans. Yet many scientists insist that Hox genes, and other transcription factors, are responsible for "laying out the flooor plan" of the organism when all they are observed to do is activate genes in already established segments of the developing embryo. We have known this for 20 years. So why are the 39 Hox genes still portrayed as determining the geometry of anatomy when all they do is bind to DNA and RNA polymerase to affect gene expression in ontogeny?
Yes, mutations in hox genes do produce aberrant body plans..like legs where there should be antennae in fruit flies. But it is a mistake to assume that this is evidence that hox genes causally lay out the body plan, just like it would be a mistake to assume that a fault in a TV, causing a disruption in the reception of the signal, shows that the TV box produces the signal itself. Linear DNA cannot produce 3D form.
I am questioning th role of Hox genes in particular, as they are the most widely touted, but I am more generallty questioning the role of transcription factors and signalling proteins as a whole since their function is only to affect the production of other proteins, not to instruct cells how to form specific 3D structures like hearts and lungs. Yes, proteins are three dimensional entities, but there is no proof that their encoded amino acid sequence alone determines their final conformation - but that is not really an issue for developmental biology. My main argument is that the body plan of the organism is not encoded in DNA, or represented in the membrane/cytoskeletal patterns of the egg. Think about it: the human brain has 10^15 synaptic connections, which is 300,000 times more than the number of base pairs in the entire genome! There is a clear difference between genomic complexity and the complexity of the organism.
Darwins doubt, pg.239
WHAT ABOUT HOX GENES?
Hox (or homeotic) genes regulate the expression of other protein-coding genes during the process of animal development. Some biologists have likened them to the conductor of an orchestra who plays the role of coordinating the contributions of the players. And because Hox genes affect so many other genes, many evo-devo advocates think that mutations in these genes can generate large- scale changes in form.
But can mutations in Hox genes transform one form of animal life—one body plan—into another? There are several reasons to doubt that they can.
First, precisely because Hox genes coordinate the expression of so many other different genes, experimentally generated mutations in Hox genes have proven harmful. in fruit flies "most mutations in homeotic [Hox] genes cause fatal birth defects." In other cases, the resulting Hox mutant phenotype, while viable in the short term, is nonetheless markedly less fit than the wild type. For example, by mutating a Hox gene in a fruit fly, biologists have produced the dramatic Antennapedia mutant, a hapless fly with legs growing out of its head where the antennae should be
Other Hox mutations have produced fruit flies in which the balancers (tiny structures behind wings that stabilize the insect in flight, called "halteres") are transformed into an extra pair of wings. Such mutations alter the structure of the animal, but not in a beneficial or permanently heritable way. The Antennapedia mutant cannot survive in the wild; it has difficulty reproducing, and its offspring die easily. Similarly, fruit-fly mutants sporting an extra set of wings lack the musculature to make use of them and, absent their balancers, cannot fly. As Hungarian evolutionary biologist Eörs Szathmáry notes with cautious understatement in the journal Nature, "macromutations of this sort [i.e., in Hox genes] are probably frequently maladaptive."
Second, Hox genes in all animal forms are expressed after the beginning of animal development, and well after the body plan has begun to be established. In fruit flies, by the time that Hox genes are expressed, roughly 6,000 cells have already formed, and the basic geometry of the fly—its anterior, posterior, dorsal, and ventral axes—is already well established. So Hox genes don't determine body-plan formation. Eric Davidson and Douglas Erwin have pointed out that Hox gene expression, although necessary for correct regional or local differentiation within a body plan, occurs much later during embryogenesis than global body-plan specification itself, which is regulated by entirely different genes. Thus, the primary origin of animal body plans in the Cambrian explosion is not merely a question of Hox gene action, but of the appearance of much deeper control elements—
Davidson's "developmental gene regulatory networks" (dGRNs).Davidson argues that it is extremely difficult to alter dGRNs without damaging their ability to regulate animal development. Third, Hox genes only provide information for building proteins that function as switches that turn other genes on and off. The genes that they regulate contain information for building proteins that form the parts of other structures and organs. The Hox genes themselves, however, do not contain information for building these structural parts. In other words, mutations in Hox genes do not have all the genetic information necessary to generate new tissues, organs, or body plans.
Nevertheless, Schwartz argues that biologists can explain complex structures such as the eye just by invoking Hox mutations alone. He asserts that "[t]here are homeobox genes for eye formation and that when one of them, the Rx gene in particular, is activated in the right place and at the right time, an individual has an eye." He also thinks that mutations in Hox genes help arrange organs to form body plans. In a review of Schwartz's book, Eörs Szathmáry finds Schwartz's reasoning deficient. He too notes that Hox genes don't code for the proteins out of which body parts are made. It follows, he insists, that mutations in Hox genes cannot by themselves build new body parts or body plans. As he explains, "Schwartz ignores the fact that homeobox genes are selector genes. They can do nothing if the genes regulated by them are not there." Though Schwartz says he has "marveled" at "the importance of homeobox genes in helping us to understand the basics of evolutionary change," kind of (nonhomologous) leg. Another homologue of the Distal-less gene in echinoderms regulates the development of tube feet and spines—anatomical features classically thought not to be homologous to arthropod limbs, nor to limbs of tetrapods.34 In each case, the Distal-less homologues play different roles determined by the higher-level organismal context. And since mutations in Hox genes do not alter higher-level epigenetic contexts,35 they cannot explain the origin of the novel epigenetic information and structure that establishes the context and that is necessary to building a new animal body plan.
Szathmáry doubts that mutations in these genes have much creative power. After asking whether Schwartz succeeds in explaining the origin of new forms of life by appealing to mutations in Hox genes, Szathmáry concludes, "I'm afraid that, in general, he does not."Nor, of course, do Hox genes possess the epigenetic information necessary for body-plan formation. Indeed, even in the best of cases mutations in Hox genes still only alter genes. Mutations in Hox genes can only generate new genetic information in DNA. They do not, and cannot, generate epigenetic information. Instead, epigenetic information and structures actually determine the function of many Hox genes, and not the reverse. This can be seen when the same Hox gene (as determined by nucleotide sequence homology) regulates the development of different anatomical features found in different phyla. For instance, in arthropods the Hox gene Distal-less is required for the normal development of jointed arthropod legs. But in vertebrates a homologous gene (e.g., the Dlx gene in mice) builds a different
Last edited by Admin on Wed Oct 28, 2015 8:20 pm; edited 2 times in total