


Convergence, another problem for evolution
https://reasonandscience.catsboard.com/t2014-convergence-another-problem-for-evolution
http://www.reasons.org/articles/convergence-evidence-for-a-single-creator
Proponents of evolution assert that convergence results when unrelated organisms encounter nearly identical selection forces (environmental, competitive, and predatory pressures). Natural selection then channels the random variations believed to be responsible for evolutionary change along similar pathways to produce similar features in unrelated organisms.
One of the challenges that convergence creates for the evolutionary paradigm is the frequency with which it occurs throughout life’s history. Convergence is a common characteristic of life. This commonness makes little sense in light of evolutionary theory. If evolution is indeed responsible for the diversity of life, one would expect convergence to be extremely rare. The mechanism that drives the evolutionary process consists of a large number of unpredictable, chance events that occur one after another. Given this mechanism and the complexity and fine-tuning of biological systems, it seems improbable that disparate evolutionary pathways would ever lead to the same biological feature.4
Two remarkable examples of complex biological features recently recognized as being convergent are bat echolocation (the ability of an organism to orient itself based on perceiving reflections of sound it emits) and parrot, songbird, and hummingbird forebrain structure. A recent DNA sequence analysis has just confirmed two earlier studies that, from an evolutionary perspective, requires echolocation in bats to have evolved independently in two separate groups (microchiroptera and megachiroptera).5-7 This study, along with previous analyses also indicate that the strikingly similar limb structures of bats and flying lemurs used for flying, likewise, must have evolved independently, when the data is interpreted from an evolutionary perspective.
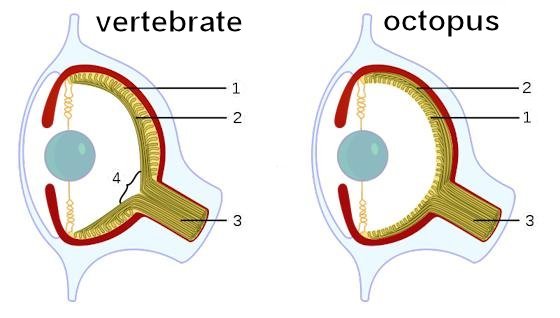
Even more challenging for the evolutionist are the cases in which convergence occurs in organisms from radically different environments. Under these circumstances, the forces that comprise natural selection must be different by definition. The classic example of this type of convergence is found in the eye structure of the cephalopods (nautili, cuttlefish, squids, and octopods).10 Their similarity to vertebrate eyes is remarkable from an evolutionary perspective, given that 1) mollusks, which include cephalopods, are classified as a member of a fundamentally different group (lophotrochozoan) than vertebrates (deuterostomes)11; and 2) the selective forces that would have shaped the formation of both the cephalopod eye and vertebrate eye must have been quite different. Evolution would have required an aquatic environment for the cephalopods and a primarily terrestrial environment for the vertebrates.
An even more remarkable example of convergence occurring in aquatic and terrestrial environments can be seen in the sandlance (fish) and chameleon (reptile), respectively. Recent experiments have uncovered an extraordinary similarity in the visual systems and behavior for these two creatures.12-15 Both the chameleon and the sandlance move their eyes independent of one another in a jerky manner, rather than in concert. While one eye is in motion the other eye is motionless. Moreover, both animals use the cornea of the eye to focus on objects. All other reptiles and fish use the lens of the eye to focus images on the retina. The chameleon and sandlance both rely on a specialized muscle (the cornealis muscle) to adjust the focusing of the cornea. The chameleon determines depth perception using a single eye. Scientists believe the sandlance also determines depth perception in this manner. Both the sandlance and the chameleon have skin coverings over their eyes to prevent them from being conspicuous to both predators and prey. The feeding behavior of both animals is also the same. The trajectory that the chameleon tongue takes when attacking its prey is the same as that taken by the sandlance when it lunges for its prey. (The sandlance buries itself in sand beds with its eyes above the surface of the sand and waits for tiny crustaceans to pass by.)
Convergence, another problem for evolution
Biologists are uncovering numerous examples of organisms that cluster together morphologically (structurally), and yet are genetically distinct. Frogs, lizards, or herbs that appear to be identical are actually different at the genetic level. An evolutionary interpretation of this data, then, demands that the morphologically identical organisms must have evolved independently of one another in a “repeatable” fashion.
“…No finale can be specified at the start, none would ever occur a second time in the same way, because any pathway proceeds through thousands of improbable stages. Alter any early event, ever so slightly, and without apparent importance at the time, and evolution cascades into a radically different channel.1
Stephen J. Gould, Wonderful Life: The Burgess Shale and the Nature of History (New York, NY: W.W. Norton & Company, 1989), 51.
Gould’s metaphor of “replaying life’s tape” asserts that if one were to push the rewind button, erase life’s history, and let the tape run again, the results would be completely different.2 The very essence of the evolutionary process renders evolutionary outcomes as nonreproducible (or nonrepeatable). Therefore, “repeatable” evolution is inconsistent with the mechanism available to bring about biological change.
Paleontologist J. William Schopf, one of the world’s leading authorities on early life on Earth, has made this very point in the book Life’s Origin.
Because biochemical systems comprise many intricately interlinked pieces, any particular full-blown system can only arise once…Since any complete biochemical system is far too elaborate to have evolved more than once in the history of life, it is safe to assume that microbes of the primal LCA cell line had the same traits that characterize all its present-day descendents.
http://www.upenn.edu/pennnews/news/evolution-unpredictable-and-irreversible-penn-biologists-show
Gould’s famous tape of life would be very different if replayed, even more different than Gould might have imagined.”
Evolutionary theorist Stephen Jay Gould is famous for describing the evolution of humans and other conscious beings as a chance accident of history. If we could go back millions of years and “run the tape of life again,” he mused, evolution would follow a different path.
Further readings :
http://www.reasons.org/articles/inability-to-repeat-the-past-dooms-evolution
http://www.reasons.org/articles/convergence-evidence-for-a-single-creator
http://www.reasons.org/blogs/the-cells-design/historical-contingency-and-the-improbability-of-protein-evolution--part-2--of-2
https://reasonandscience.catsboard.com/t2014-convergence-another-problem-for-evolution
http://www.reasons.org/articles/convergence-evidence-for-a-single-creator
Proponents of evolution assert that convergence results when unrelated organisms encounter nearly identical selection forces (environmental, competitive, and predatory pressures). Natural selection then channels the random variations believed to be responsible for evolutionary change along similar pathways to produce similar features in unrelated organisms.
One of the challenges that convergence creates for the evolutionary paradigm is the frequency with which it occurs throughout life’s history. Convergence is a common characteristic of life. This commonness makes little sense in light of evolutionary theory. If evolution is indeed responsible for the diversity of life, one would expect convergence to be extremely rare. The mechanism that drives the evolutionary process consists of a large number of unpredictable, chance events that occur one after another. Given this mechanism and the complexity and fine-tuning of biological systems, it seems improbable that disparate evolutionary pathways would ever lead to the same biological feature.4
Two remarkable examples of complex biological features recently recognized as being convergent are bat echolocation (the ability of an organism to orient itself based on perceiving reflections of sound it emits) and parrot, songbird, and hummingbird forebrain structure. A recent DNA sequence analysis has just confirmed two earlier studies that, from an evolutionary perspective, requires echolocation in bats to have evolved independently in two separate groups (microchiroptera and megachiroptera).5-7 This study, along with previous analyses also indicate that the strikingly similar limb structures of bats and flying lemurs used for flying, likewise, must have evolved independently, when the data is interpreted from an evolutionary perspective.
Even more challenging for the evolutionist are the cases in which convergence occurs in organisms from radically different environments. Under these circumstances, the forces that comprise natural selection must be different by definition. The classic example of this type of convergence is found in the eye structure of the cephalopods (nautili, cuttlefish, squids, and octopods).10 Their similarity to vertebrate eyes is remarkable from an evolutionary perspective, given that 1) mollusks, which include cephalopods, are classified as a member of a fundamentally different group (lophotrochozoan) than vertebrates (deuterostomes)11; and 2) the selective forces that would have shaped the formation of both the cephalopod eye and vertebrate eye must have been quite different. Evolution would have required an aquatic environment for the cephalopods and a primarily terrestrial environment for the vertebrates.
An even more remarkable example of convergence occurring in aquatic and terrestrial environments can be seen in the sandlance (fish) and chameleon (reptile), respectively. Recent experiments have uncovered an extraordinary similarity in the visual systems and behavior for these two creatures.12-15 Both the chameleon and the sandlance move their eyes independent of one another in a jerky manner, rather than in concert. While one eye is in motion the other eye is motionless. Moreover, both animals use the cornea of the eye to focus on objects. All other reptiles and fish use the lens of the eye to focus images on the retina. The chameleon and sandlance both rely on a specialized muscle (the cornealis muscle) to adjust the focusing of the cornea. The chameleon determines depth perception using a single eye. Scientists believe the sandlance also determines depth perception in this manner. Both the sandlance and the chameleon have skin coverings over their eyes to prevent them from being conspicuous to both predators and prey. The feeding behavior of both animals is also the same. The trajectory that the chameleon tongue takes when attacking its prey is the same as that taken by the sandlance when it lunges for its prey. (The sandlance buries itself in sand beds with its eyes above the surface of the sand and waits for tiny crustaceans to pass by.)
Convergence, another problem for evolution
Biologists are uncovering numerous examples of organisms that cluster together morphologically (structurally), and yet are genetically distinct. Frogs, lizards, or herbs that appear to be identical are actually different at the genetic level. An evolutionary interpretation of this data, then, demands that the morphologically identical organisms must have evolved independently of one another in a “repeatable” fashion.
“…No finale can be specified at the start, none would ever occur a second time in the same way, because any pathway proceeds through thousands of improbable stages. Alter any early event, ever so slightly, and without apparent importance at the time, and evolution cascades into a radically different channel.1
Stephen J. Gould, Wonderful Life: The Burgess Shale and the Nature of History (New York, NY: W.W. Norton & Company, 1989), 51.
Gould’s metaphor of “replaying life’s tape” asserts that if one were to push the rewind button, erase life’s history, and let the tape run again, the results would be completely different.2 The very essence of the evolutionary process renders evolutionary outcomes as nonreproducible (or nonrepeatable). Therefore, “repeatable” evolution is inconsistent with the mechanism available to bring about biological change.
Paleontologist J. William Schopf, one of the world’s leading authorities on early life on Earth, has made this very point in the book Life’s Origin.
Because biochemical systems comprise many intricately interlinked pieces, any particular full-blown system can only arise once…Since any complete biochemical system is far too elaborate to have evolved more than once in the history of life, it is safe to assume that microbes of the primal LCA cell line had the same traits that characterize all its present-day descendents.
http://www.upenn.edu/pennnews/news/evolution-unpredictable-and-irreversible-penn-biologists-show
Gould’s famous tape of life would be very different if replayed, even more different than Gould might have imagined.”
Evolutionary theorist Stephen Jay Gould is famous for describing the evolution of humans and other conscious beings as a chance accident of history. If we could go back millions of years and “run the tape of life again,” he mused, evolution would follow a different path.
Further readings :
http://www.reasons.org/articles/inability-to-repeat-the-past-dooms-evolution
http://www.reasons.org/articles/convergence-evidence-for-a-single-creator
http://www.reasons.org/blogs/the-cells-design/historical-contingency-and-the-improbability-of-protein-evolution--part-2--of-2
Last edited by Admin on Sat Apr 25, 2020 1:47 am; edited 7 times in total






 This unique strategy has evolved in cone shells Conus, of which there are some 500-600 world species. Cone shells are carnivores that employ modified radular cusps or teeth as harpoons (see photograph below and in situ on Right) to stun polychaete, snail and, in some cases, fish prey, before feeding on them. Cone shells lack a "traditional" scraping radula. Prey if small are swallowed whole by drawing them via the "
This unique strategy has evolved in cone shells Conus, of which there are some 500-600 world species. Cone shells are carnivores that employ modified radular cusps or teeth as harpoons (see photograph below and in situ on Right) to stun polychaete, snail and, in some cases, fish prey, before feeding on them. Cone shells lack a "traditional" scraping radula. Prey if small are swallowed whole by drawing them via the " The teeth or harpoons are constructed of a flat sheet of chitin and protein rolled up to enclose a spiral channel or duct that may be only partially closed off from the outside. The teeth are secreted in the long end of a Y- or L-shaped radula sac and stored in the short end, oriented there much arrows in a quiver mostly with their pointy ends facing towards the opening of the storage sac into the pharynx (see drawing on Right). The teeth are hardened by quinone-tanning and by inclusion of calcium. They are moved individually as needed to the end of the proboscis where they are firmly held by a special sphincter muscle. While no-one seems to know exactly how they get from the storage sac to the tip of the proboscis, one idea is that the proboscis inverts on itself, and in this inside-out form reaches back to envelope a tooth completely. Then, gripping the basal end of tooth, the proboscis everts with the harpoon now pointing in the correct direction from the tip of the proboscis. Simpler still might be for the tooth to be just moved into position through muscular paristalses of the proboscis from the storage sac into the pharynx and then into the proboscis (see drawing on Right).
The teeth or harpoons are constructed of a flat sheet of chitin and protein rolled up to enclose a spiral channel or duct that may be only partially closed off from the outside. The teeth are secreted in the long end of a Y- or L-shaped radula sac and stored in the short end, oriented there much arrows in a quiver mostly with their pointy ends facing towards the opening of the storage sac into the pharynx (see drawing on Right). The teeth are hardened by quinone-tanning and by inclusion of calcium. They are moved individually as needed to the end of the proboscis where they are firmly held by a special sphincter muscle. While no-one seems to know exactly how they get from the storage sac to the tip of the proboscis, one idea is that the proboscis inverts on itself, and in this inside-out form reaches back to envelope a tooth completely. Then, gripping the basal end of tooth, the proboscis everts with the harpoon now pointing in the correct direction from the tip of the proboscis. Simpler still might be for the tooth to be just moved into position through muscular paristalses of the proboscis from the storage sac into the pharynx and then into the proboscis (see drawing on Right).