One Way To Think About the Complexity of the “Simplest” Life Form 1
I have always been fascinated by the question, “How simple can life get?” After all, anything that is alive has to perform certain functions such as reacting to external stimuli, taking in energy and converting that energy to its own use, reproducing, etc. Exactly how simple can a living system be if it has to perform such tasks? Many biologists have investigated this question, but there isn’t a firm answer. Typically, biologists talk about how simple a genome can be. The simplest genome belongs to a bacterium known as Carsonella ruddii. It has 159,662 base pairs in its genome, which is thought to contain 182 genes.1 However, it is not considered a real living organism, as it cannot perform all the functions of life without the help of cells found in jumping plant lice.
The bacterium known as Pelagibacter ubique has the smallest genome of any truly free-living organism. It weighs in at 1,308,759 base pairs and 1,354 genes.2 However, there is something in between these two bacteria that might qualify as a real living organism. It is the bacterium Mycoplasma genitalium. It’s genome has 582,970 base pairs and 525 genes.3 While it is a parasite, it performs all the standard functions of life on its own. It just uses other organisms (people as well as animals of the order Primates) for food and housing. Thus, while it cannot exist without other organisms, it might be the best indicator of how “simple” life can get.
If you follow science news at all, you might recognize the name. Two years ago, Dr Craig Venter and his team constructed their own version of that bacterium with the help of living versions of the bacterium, yeast cells, and bacteria of another species from the same genus. Well, now a scientist from Venter’s lab teamed up with several scientists from Stanford University to produce a computer simulation of the bacterium!
Their work, which seems truly marvelous, gives us deep insight into how complex the “simplest” living organism really is.
Let’s start with what the computer simulation actually accomplished. It modeled all the inputs and outputs of the bacterium’s 525 genes throughout a single cell cycle. In other words, it simulated how the genome produces proteins, how those proteins interact with other proteins, and how the entire system is regulated. It followed these processes through all the events leading up to and including the cell reproducing itself.4
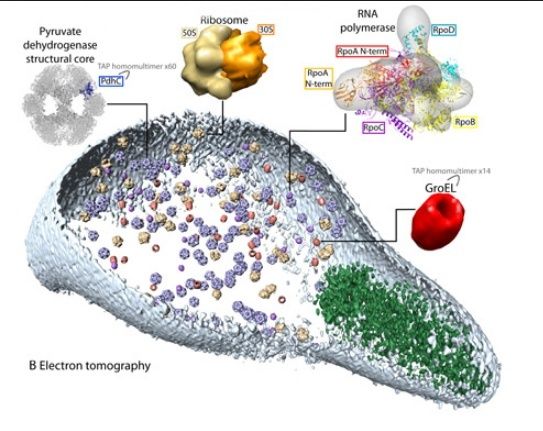
Now that’s a lot of work! How did the authors do it? Well, they looked at over 900 different scientific papers that had been produced on the inner workings of Mycoplasma genitalium, and they identified 1,900 specific parameters that seem to govern how the cell operates. There were several discrepancies that were found among the papers involved, and as a result, there was a lot of reconciliation that had to be done. The details of this reconciliation and other matters are found in a 120-page supplement to the 12-page scientific paper.
Once the reconciliation of these studies was accomplished, the essential workings of the cell were split into 28 separate modules that each governed specific functions of the cell. For example, one module dealt with metabolism, while another dealt with the activation of proteins once they were produced. Once each module was built and tested individually, the modules were then joined by looking at what they produced every second. If the products of one module were the kinds of chemicals used by a second module, those products were then treated as inputs to the second module for the next second of computation. The computation proceeded like this (checking the inputs and outputs of each module) for about 10 hours, which is roughly the time it takes a real Mycoplasma genitalium to reproduce.
Why would a group want to undertake such a complex endeavor? Well, one obvious reason is the reconciliation that I mentioned previously. As independent papers, each of the 900 studies to which the authors referred made sense. However, when the authors started using the results of those studies in a model that tries to take all the molecular processes of a cell into account, they found that some results didn’t mesh well with others. The reconciliation that had to take place to get the simulation working will help us better understand the limits of many of the studies related to Mycoplasma genitalium and hopefully will lead to more detailed studies that will slowly wipe away such discrepancies. Also, as the authors state, these kinds of models will:
…accelerate biological discovery and bioengineering by facilitating experimental design and interpretation. Morever, [this study and others] raise the exciting possibility of using whole-cell models to enable computer-aided rational design of novel microorganisms.
So in the end, not only will such models help us better design and interpret experiments, they might one day lead us to ways that we can engineer new microorganisms.
This is fantastic work, and I do think it opens up new vistas in cell and molecular biology. However, we need to pull back for a moment and think about the direct implications of this computer simulation. It simulated, in very basic terms, the molecular interactions that occur in a cell that might be a good analog for the simplest possible life form. It skipped over a lot of details, of course, so it is not a complete simulation by any means. Nevertheless, it is a great first step towards understanding how a living system really works.
Now let’s look at this in very practical terms. In order to be able to match the speed at which the organism operates, this less-than-complete simulation required a cluster of 128 computers to get the job done. Think about that for a moment. In order to simulate most (but not all) of the processes that take place in an analog for what might be the simplest possible living organism, the authors needed the power of 128 computers running together! That should tell us something very clearly:
There is no such thing as a simple living organism.
The more we understand life, the more clear it becomes that even the “simplest” version of it has to be the result of design.
REFERENCES
1. Atsushi Nakabachi, et al., “The 160-Kilobase Genome of the Bacterial Endosymbiont Carsonella,” Science 314:267, 2006.
Return to Text
2. Stephen J. Giovannoni, et al., “Genome Streamlining in a Cosmopolitan Oceanic Bacterium,” Science 309:1242-1245, 2005.
Return to Text
3. According to the Comprehensive Microbial Resource Manual.
Return to Text
4. Jonathan R. Karr, et al., “A Whole-Cell Computational Model Predicts Phenotype from Genotype,” Cell 150(2):389-401, 2012.
Return to Text
1) http://blog.drwile.com/?p=8161
I have always been fascinated by the question, “How simple can life get?” After all, anything that is alive has to perform certain functions such as reacting to external stimuli, taking in energy and converting that energy to its own use, reproducing, etc. Exactly how simple can a living system be if it has to perform such tasks? Many biologists have investigated this question, but there isn’t a firm answer. Typically, biologists talk about how simple a genome can be. The simplest genome belongs to a bacterium known as Carsonella ruddii. It has 159,662 base pairs in its genome, which is thought to contain 182 genes.1 However, it is not considered a real living organism, as it cannot perform all the functions of life without the help of cells found in jumping plant lice.
The bacterium known as Pelagibacter ubique has the smallest genome of any truly free-living organism. It weighs in at 1,308,759 base pairs and 1,354 genes.2 However, there is something in between these two bacteria that might qualify as a real living organism. It is the bacterium Mycoplasma genitalium. It’s genome has 582,970 base pairs and 525 genes.3 While it is a parasite, it performs all the standard functions of life on its own. It just uses other organisms (people as well as animals of the order Primates) for food and housing. Thus, while it cannot exist without other organisms, it might be the best indicator of how “simple” life can get.
If you follow science news at all, you might recognize the name. Two years ago, Dr Craig Venter and his team constructed their own version of that bacterium with the help of living versions of the bacterium, yeast cells, and bacteria of another species from the same genus. Well, now a scientist from Venter’s lab teamed up with several scientists from Stanford University to produce a computer simulation of the bacterium!
Their work, which seems truly marvelous, gives us deep insight into how complex the “simplest” living organism really is.
Let’s start with what the computer simulation actually accomplished. It modeled all the inputs and outputs of the bacterium’s 525 genes throughout a single cell cycle. In other words, it simulated how the genome produces proteins, how those proteins interact with other proteins, and how the entire system is regulated. It followed these processes through all the events leading up to and including the cell reproducing itself.4
Now that’s a lot of work! How did the authors do it? Well, they looked at over 900 different scientific papers that had been produced on the inner workings of Mycoplasma genitalium, and they identified 1,900 specific parameters that seem to govern how the cell operates. There were several discrepancies that were found among the papers involved, and as a result, there was a lot of reconciliation that had to be done. The details of this reconciliation and other matters are found in a 120-page supplement to the 12-page scientific paper.
Once the reconciliation of these studies was accomplished, the essential workings of the cell were split into 28 separate modules that each governed specific functions of the cell. For example, one module dealt with metabolism, while another dealt with the activation of proteins once they were produced. Once each module was built and tested individually, the modules were then joined by looking at what they produced every second. If the products of one module were the kinds of chemicals used by a second module, those products were then treated as inputs to the second module for the next second of computation. The computation proceeded like this (checking the inputs and outputs of each module) for about 10 hours, which is roughly the time it takes a real Mycoplasma genitalium to reproduce.
Why would a group want to undertake such a complex endeavor? Well, one obvious reason is the reconciliation that I mentioned previously. As independent papers, each of the 900 studies to which the authors referred made sense. However, when the authors started using the results of those studies in a model that tries to take all the molecular processes of a cell into account, they found that some results didn’t mesh well with others. The reconciliation that had to take place to get the simulation working will help us better understand the limits of many of the studies related to Mycoplasma genitalium and hopefully will lead to more detailed studies that will slowly wipe away such discrepancies. Also, as the authors state, these kinds of models will:
…accelerate biological discovery and bioengineering by facilitating experimental design and interpretation. Morever, [this study and others] raise the exciting possibility of using whole-cell models to enable computer-aided rational design of novel microorganisms.
So in the end, not only will such models help us better design and interpret experiments, they might one day lead us to ways that we can engineer new microorganisms.
This is fantastic work, and I do think it opens up new vistas in cell and molecular biology. However, we need to pull back for a moment and think about the direct implications of this computer simulation. It simulated, in very basic terms, the molecular interactions that occur in a cell that might be a good analog for the simplest possible life form. It skipped over a lot of details, of course, so it is not a complete simulation by any means. Nevertheless, it is a great first step towards understanding how a living system really works.
Now let’s look at this in very practical terms. In order to be able to match the speed at which the organism operates, this less-than-complete simulation required a cluster of 128 computers to get the job done. Think about that for a moment. In order to simulate most (but not all) of the processes that take place in an analog for what might be the simplest possible living organism, the authors needed the power of 128 computers running together! That should tell us something very clearly:
There is no such thing as a simple living organism.
The more we understand life, the more clear it becomes that even the “simplest” version of it has to be the result of design.
REFERENCES
1. Atsushi Nakabachi, et al., “The 160-Kilobase Genome of the Bacterial Endosymbiont Carsonella,” Science 314:267, 2006.
Return to Text
2. Stephen J. Giovannoni, et al., “Genome Streamlining in a Cosmopolitan Oceanic Bacterium,” Science 309:1242-1245, 2005.
Return to Text
3. According to the Comprehensive Microbial Resource Manual.
Return to Text
4. Jonathan R. Karr, et al., “A Whole-Cell Computational Model Predicts Phenotype from Genotype,” Cell 150(2):389-401, 2012.
Return to Text
1) http://blog.drwile.com/?p=8161
Last edited by Admin on Wed Jul 22, 2015 1:25 pm; edited 2 times in total