


Wanna Build a Cell? A DVD Player Might Be Easier 1
https://reasonandscience.catsboard.com/t2404-wanna-build-a-cell-a-dvd-player-might-be-easier
Wanna Build a Cell? A DVD Player Might Be Easier Imagine that you’re building the world’s first DVD player. What must you have before you can turn it on and watch a movie for the first time?
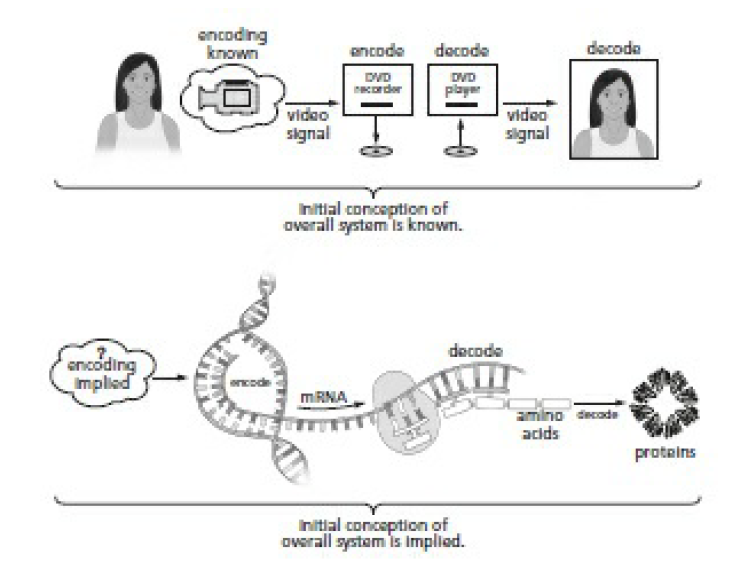
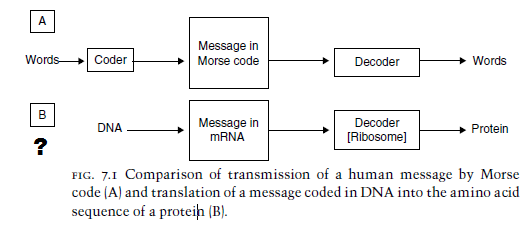
A DVD. How do you get a DVD? You need a DVD recorder first. How do you make a DVD recorder? First you have to define the language. When Russell Kirsch (who we met in chapter Cool created the world’s first digital image, he had to define a language for images first. Likewise you have to define the language that gets written on the DVD, then build hardware that speaks that language. Language must be defined first. Our DVD recorder/player problem is an encoding-decoding problem, just like the information in DNA. You’ll recall that communication, by definition, requires four things to exist:
1. A code
2. An encoder that obeys the rules of a code
3. A message that obeys the rules of the code
4. A decoder that obeys the rules of the code
These four things—language, transmitter of language, message, and receiver of language—all have to be precisely defined in advance before any form of communication can be possible at all.
A camera sends a signal to a DVD recorder, which records a DVD. The DVD player reads the DVD and converts it to a TV signal. This is conceptually identical to DNA translation. The only difference is that we don’t know how the original signal—the pattern in the first DNA strand—was encoded. The first DNA strand had to contain a plan to build something, and that plan had to get there somehow. An original encoder that translates the idea of an organism into instructions to build the organism (analogous to the camera) is directly implied.
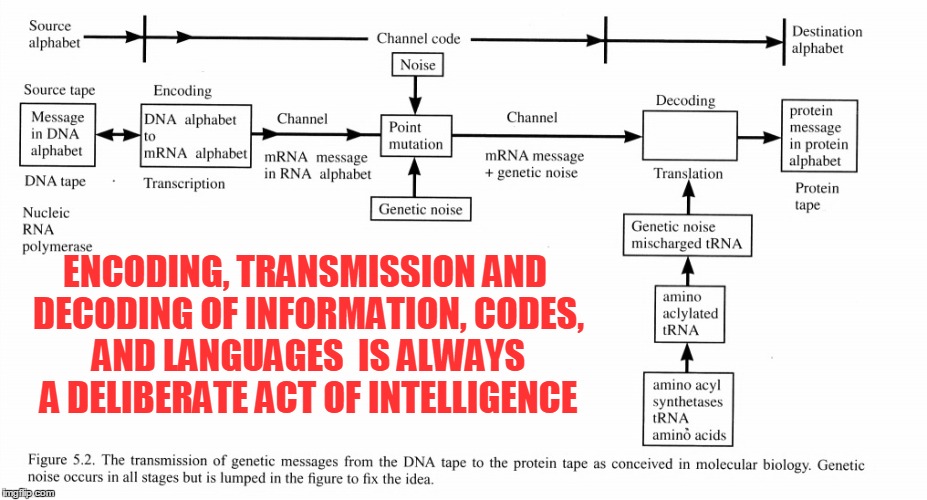
The rules of any communication system are always defined in advance by a process of deliberate choices. There must be prearranged agreement between sender and receiver, otherwise communication is impossible. By definition, a communication system cannot evolve from something simpler because evolution itself requires communication to exist first. You can’t make copies of a message without the message, and you can’t create a message without first having a language. And before that, you need intent. A code is an abstract, immaterial, nonphysical set of rules. There is no physical law that says ink on a piece of paper formed in the shape T-R-E-E should correspond to that large leafy organism in your front yard. You cannot derive the local rules of a code from the laws of physics, because hard physical laws necessarily exclude choice. On the other hand, the coder decides whether “1” means “on” or “off.” She decides whether “0” means “off” or “on.” Codes, by definition, are freely chosen. The rules of the code come before all else. These rules of any language are chosen with a goal in mind: communication, which is always driven by intent. That being said, conscious beings can evolve a simple code into a more complex code—if they can communicate in the first place. But even simple grunts and hand motions between two humans who share no language still require communication to occur. Pointing to a table and making a sound that means “table” still requires someone to recognize what your pointing finger means.
A number of special natural laws can be derived from general natural law NLI-4: 2
- NLI-4a: Every code is based on a mutual agreement between sender and receiver.
- NLI-4b: There can be no new information without an intelligent sender.
- NLI-4c: Every information transmission chain can be traced back to an intelligent sender.
- NLI-4d: The assignment of meaning to a set of symbols is a mental process requiring intelligence.
According to NLI-4, there is an intelligent originator at the beginning of every information transmission chain. If this principle is applied consistently to biological information, an intelligent originator is necessary here as well.
DNA Is a Structure That Encodes Biological Information
What do a human, a rose, and a bacterium have in common? Each of these things — along with every other organism on Earth — contains the molecular instructions for life, called deoxyribonucleic acid or DNA. Encoded within this DNA are the directions for traits as diverse as the color of a person's eyes, the scent of a rose, and the way in which bacteria infect a lung cell.
http://www.nature.com/scitable/topicpage/dna-is-a-structure-that-encodes-biological-6493050
Transmission of intra-cellular genetic information: A system proposal
The model of a transmission system of genetic information is concerned with the identification, reproduction and mathematical classification of the nucleotide sequence of single stranded DNA by the genetic encoder. Hence, a genetic encoder is devised where labelings and cyclic codes are established. The establishment of the algebraic structure of the corresponding codes alphabets, mappings, labelings, primitive polynomials (p(x)) and code generator polynomials (g(x )) are quite important in characterizing error-correcting codes subclasses of G-linear codes.
http://www.sciencedirect.com/science/article/pii/S0022519314003233
The Information in DNA Is Decoded by Transcription
DNA is essentially a storage molecule. It contains all of the instructions a cell needs to sustain itself. These instructions are found within genes, which are sections of DNA made up of specific sequences of nucleotides. In order to be implemented, the instructions contained within genes must be expressed, or copied into a form that can be used by cells to produce the proteins needed to support life.
The instructions stored within DNA are read and processed by a cell in two steps: transcription and translation. Each of these steps is a separate biochemical process involving multiple molecules. During transcription, a portion of the cell's DNA serves as a template for creation of an RNA molecule. (RNA, or ribonucleic acid, is chemically similar to DNA, except for three main differences described later on in this concept page.) In some cases, the newly created RNA molecule is itself a finished product, and it serves an important function within the cell. In other cases, the RNA molecule carries messages from the DNA to other parts of the cell for processing. Most often, this information is used to manufacture proteins. The specific type of RNA that carries the information stored in DNA to other areas of the cell is called messenger RNA, or mRNA.
http://www.nature.com/scitable/topicpage/the-information-in-dna-is-decoded-by-6524808
1. Perry Marshall, Evolution 2.0, page 153
2. http://www.0095.info/en/
More information : The genetic code, insurmountable problem for non-intelligent origin
http://reasonandscience.heavenforum.org/t2363-the-genetic-code-unsurmountable-problem-for-non-intelligent-origin
https://reasonandscience.catsboard.com/t2404-wanna-build-a-cell-a-dvd-player-might-be-easier
Wanna Build a Cell? A DVD Player Might Be Easier Imagine that you’re building the world’s first DVD player. What must you have before you can turn it on and watch a movie for the first time?
A DVD. How do you get a DVD? You need a DVD recorder first. How do you make a DVD recorder? First you have to define the language. When Russell Kirsch (who we met in chapter Cool created the world’s first digital image, he had to define a language for images first. Likewise you have to define the language that gets written on the DVD, then build hardware that speaks that language. Language must be defined first. Our DVD recorder/player problem is an encoding-decoding problem, just like the information in DNA. You’ll recall that communication, by definition, requires four things to exist:
1. A code
2. An encoder that obeys the rules of a code
3. A message that obeys the rules of the code
4. A decoder that obeys the rules of the code
These four things—language, transmitter of language, message, and receiver of language—all have to be precisely defined in advance before any form of communication can be possible at all.
A camera sends a signal to a DVD recorder, which records a DVD. The DVD player reads the DVD and converts it to a TV signal. This is conceptually identical to DNA translation. The only difference is that we don’t know how the original signal—the pattern in the first DNA strand—was encoded. The first DNA strand had to contain a plan to build something, and that plan had to get there somehow. An original encoder that translates the idea of an organism into instructions to build the organism (analogous to the camera) is directly implied.
The rules of any communication system are always defined in advance by a process of deliberate choices. There must be prearranged agreement between sender and receiver, otherwise communication is impossible. By definition, a communication system cannot evolve from something simpler because evolution itself requires communication to exist first. You can’t make copies of a message without the message, and you can’t create a message without first having a language. And before that, you need intent. A code is an abstract, immaterial, nonphysical set of rules. There is no physical law that says ink on a piece of paper formed in the shape T-R-E-E should correspond to that large leafy organism in your front yard. You cannot derive the local rules of a code from the laws of physics, because hard physical laws necessarily exclude choice. On the other hand, the coder decides whether “1” means “on” or “off.” She decides whether “0” means “off” or “on.” Codes, by definition, are freely chosen. The rules of the code come before all else. These rules of any language are chosen with a goal in mind: communication, which is always driven by intent. That being said, conscious beings can evolve a simple code into a more complex code—if they can communicate in the first place. But even simple grunts and hand motions between two humans who share no language still require communication to occur. Pointing to a table and making a sound that means “table” still requires someone to recognize what your pointing finger means.
A number of special natural laws can be derived from general natural law NLI-4: 2
- NLI-4a: Every code is based on a mutual agreement between sender and receiver.
- NLI-4b: There can be no new information without an intelligent sender.
- NLI-4c: Every information transmission chain can be traced back to an intelligent sender.
- NLI-4d: The assignment of meaning to a set of symbols is a mental process requiring intelligence.
According to NLI-4, there is an intelligent originator at the beginning of every information transmission chain. If this principle is applied consistently to biological information, an intelligent originator is necessary here as well.
DNA Is a Structure That Encodes Biological Information
What do a human, a rose, and a bacterium have in common? Each of these things — along with every other organism on Earth — contains the molecular instructions for life, called deoxyribonucleic acid or DNA. Encoded within this DNA are the directions for traits as diverse as the color of a person's eyes, the scent of a rose, and the way in which bacteria infect a lung cell.
http://www.nature.com/scitable/topicpage/dna-is-a-structure-that-encodes-biological-6493050
Transmission of intra-cellular genetic information: A system proposal
The model of a transmission system of genetic information is concerned with the identification, reproduction and mathematical classification of the nucleotide sequence of single stranded DNA by the genetic encoder. Hence, a genetic encoder is devised where labelings and cyclic codes are established. The establishment of the algebraic structure of the corresponding codes alphabets, mappings, labelings, primitive polynomials (p(x)) and code generator polynomials (g(x )) are quite important in characterizing error-correcting codes subclasses of G-linear codes.
http://www.sciencedirect.com/science/article/pii/S0022519314003233
The Information in DNA Is Decoded by Transcription
DNA is essentially a storage molecule. It contains all of the instructions a cell needs to sustain itself. These instructions are found within genes, which are sections of DNA made up of specific sequences of nucleotides. In order to be implemented, the instructions contained within genes must be expressed, or copied into a form that can be used by cells to produce the proteins needed to support life.
The instructions stored within DNA are read and processed by a cell in two steps: transcription and translation. Each of these steps is a separate biochemical process involving multiple molecules. During transcription, a portion of the cell's DNA serves as a template for creation of an RNA molecule. (RNA, or ribonucleic acid, is chemically similar to DNA, except for three main differences described later on in this concept page.) In some cases, the newly created RNA molecule is itself a finished product, and it serves an important function within the cell. In other cases, the RNA molecule carries messages from the DNA to other parts of the cell for processing. Most often, this information is used to manufacture proteins. The specific type of RNA that carries the information stored in DNA to other areas of the cell is called messenger RNA, or mRNA.
http://www.nature.com/scitable/topicpage/the-information-in-dna-is-decoded-by-6524808
1. Perry Marshall, Evolution 2.0, page 153
2. http://www.0095.info/en/
More information : The genetic code, insurmountable problem for non-intelligent origin
http://reasonandscience.heavenforum.org/t2363-the-genetic-code-unsurmountable-problem-for-non-intelligent-origin
Last edited by Admin on Sun 15 Jul 2018 - 4:24; edited 7 times in total