Centriole/Centrosome: The centriole spindle the most complex machine known in nature
Centrosome 1

In cell biology, the centrosome (Latin centrum 'center' + Greek sōma 'body') is an organelle that serves as the main microtubule organizing center (MTOC) of the animal cell as well as a regulator of cell-cycle progression. The centrosome is thought to have evolved only in the metazoan lineage of eukaryotic cells. Fungi and plants lack centrosomes and therefore use other MTOC structures to organize their microtubules. Although the centrosome has a key role in efficient mitosis in animal cells, it is not essential.
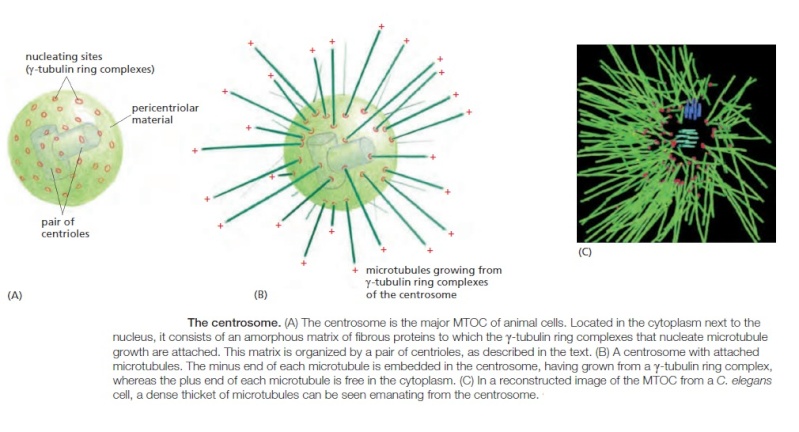
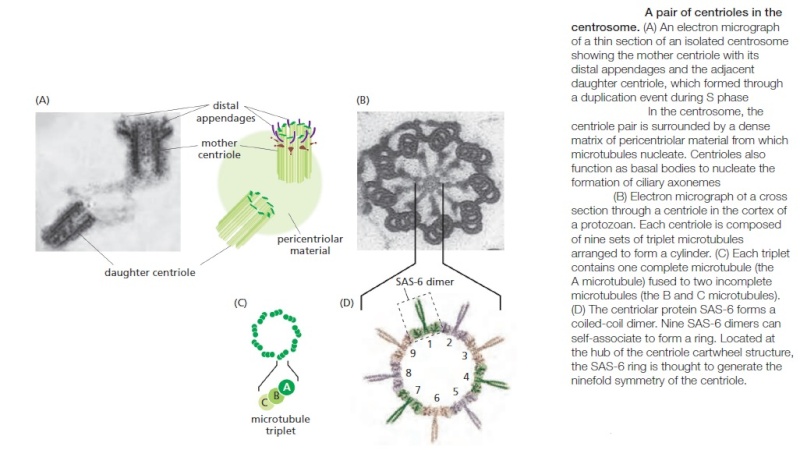
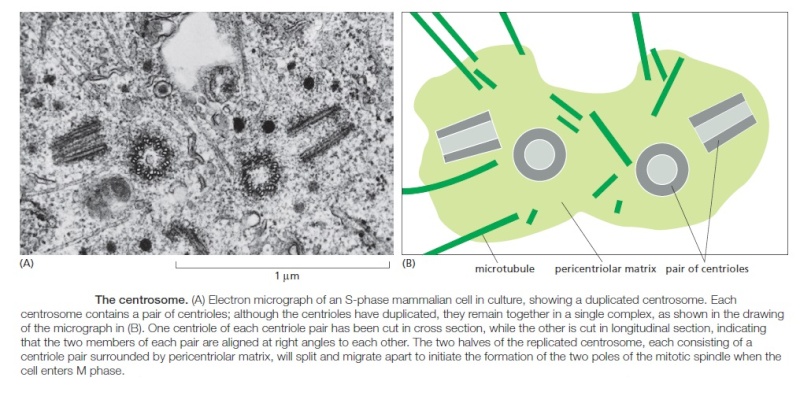
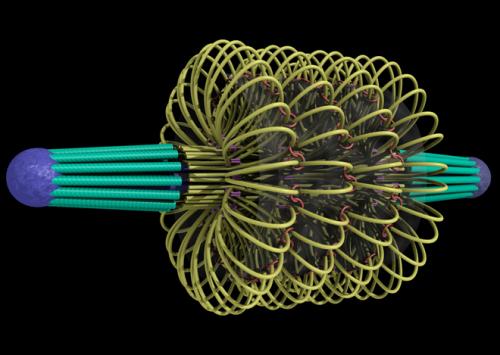
Centrosomes are composed of two orthogonally arranged centrioles surrounded by an amorphous mass of protein termed the pericentriolar material (PCM). The PCM contains proteins responsible for microtubule nucleation and anchoring including γ-tubulin, pericentrin and ninein. In general, each centriole of the centrosome is based on a nine triplet microtubule assembled in a cartwheel structure, and contains centrin, cenexin and tektin.
Functions
Role of the centrosome in cell cycle progression
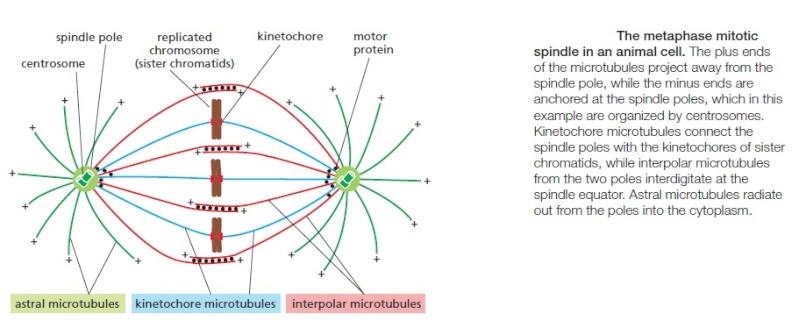
Centrosomes are associated with the nuclear membrane during prophase of the cell cycle. In mitosis the nuclear membrane breaks down and the centrosome nucleated microtubules (parts of the cytoskeleton) can interact with the chromosomes to build the mitotic spindle.
The mother centriole, the older of the two in the centriole pair, also has a central role in making cilia and flagella.
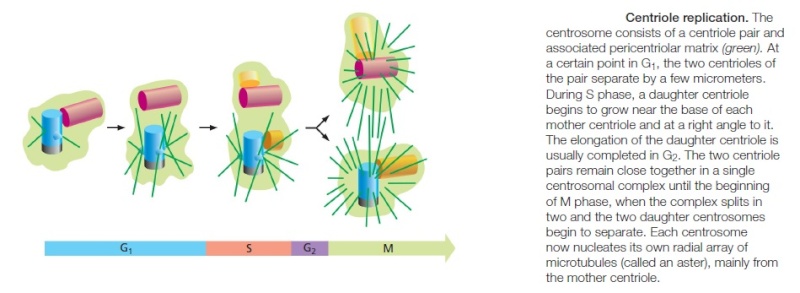
The centrosome is copied only once per cell cycle so that each daughter cell inherits one centrosome, containing two structures called centrioles

Centrosome Duplication
Cell Cycle Regulation of Centrosome Duplication
Centrosome duplication is heavily regulated by cell cycle controls. This link between the cell cycle and the centrosome cycle is mediated by cyclin-dependent kinase 2 (Cdk2). There has been ample evidence that Cdk2 is necessary for both DNA replication and centrosome duplication, which are both key events in S phase. It has also been shown that Cdk2 complexes with both cyclin A and cyclin E and this complex is critical for centrosome duplication. Three Cdk2 substrates have been proposed to be responsible for regulation of centriole duplication. These include: nucleophosmin (NPM/B23), CP110, and MPS1.[3] Nucleophosmin is only found in unreplicated centrosomes and it’s phosphorylation by Cdk2/cyclin E removes NPM from the centrosomes, initiating procentriole formation. CP110 is an important centrosomal protein that is phosphorylated by both mitotic and interphase Cdk/cyclin complexes and is thought to influence centrosome duplication in S phase. MPS1 is a protein kinase that is essential to the spindle assembly checkpoint, and may remodel an SAS6-cored intermediate between severed mother and daughter centrioles into a pair of cartwheel protein complexes onto which procentrioles assemble.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3947860/
Centrosome 1

In cell biology, the centrosome (Latin centrum 'center' + Greek sōma 'body') is an organelle that serves as the main microtubule organizing center (MTOC) of the animal cell as well as a regulator of cell-cycle progression. The centrosome is thought to have evolved only in the metazoan lineage of eukaryotic cells. Fungi and plants lack centrosomes and therefore use other MTOC structures to organize their microtubules. Although the centrosome has a key role in efficient mitosis in animal cells, it is not essential.
Centrosomes are composed of two orthogonally arranged centrioles surrounded by an amorphous mass of protein termed the pericentriolar material (PCM). The PCM contains proteins responsible for microtubule nucleation and anchoring including γ-tubulin, pericentrin and ninein. In general, each centriole of the centrosome is based on a nine triplet microtubule assembled in a cartwheel structure, and contains centrin, cenexin and tektin.
Functions
Role of the centrosome in cell cycle progression
Centrosomes are associated with the nuclear membrane during prophase of the cell cycle. In mitosis the nuclear membrane breaks down and the centrosome nucleated microtubules (parts of the cytoskeleton) can interact with the chromosomes to build the mitotic spindle.
The mother centriole, the older of the two in the centriole pair, also has a central role in making cilia and flagella.
The centrosome is copied only once per cell cycle so that each daughter cell inherits one centrosome, containing two structures called centrioles
Centrosome Duplication
Cell Cycle Regulation of Centrosome Duplication
Centrosome duplication is heavily regulated by cell cycle controls. This link between the cell cycle and the centrosome cycle is mediated by cyclin-dependent kinase 2 (Cdk2). There has been ample evidence that Cdk2 is necessary for both DNA replication and centrosome duplication, which are both key events in S phase. It has also been shown that Cdk2 complexes with both cyclin A and cyclin E and this complex is critical for centrosome duplication. Three Cdk2 substrates have been proposed to be responsible for regulation of centriole duplication. These include: nucleophosmin (NPM/B23), CP110, and MPS1.[3] Nucleophosmin is only found in unreplicated centrosomes and it’s phosphorylation by Cdk2/cyclin E removes NPM from the centrosomes, initiating procentriole formation. CP110 is an important centrosomal protein that is phosphorylated by both mitotic and interphase Cdk/cyclin complexes and is thought to influence centrosome duplication in S phase. MPS1 is a protein kinase that is essential to the spindle assembly checkpoint, and may remodel an SAS6-cored intermediate between severed mother and daughter centrioles into a pair of cartwheel protein complexes onto which procentrioles assemble.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3947860/
Last edited by Admin on Wed Aug 29, 2018 2:27 am; edited 2 times in total