Centriole biogenesis, and the duplication cycle, amazing evidence of design
https://reasonandscience.catsboard.com/t2090-centriole-biogenesis-and-the-duplication-cycle-amazing-evidence-of-design
The duplication of eukaryotic cells is a all fine-tuned biochemical processes that depends on the precise structural arrangement of the cellular components. Mitotic cell division is the most fundamental task of all living cells. Cells have intricate and tightly regulated machinery to ensure that mitosis occurs with appropriate frequency and high fidelity.
The only way to make a new cell is to duplicate a cell that already exists. A cell reproduces by performing an orderly sequence of events in which it duplicates its contents and then divides in two. This cycle of duplication and division, known as the cell cycle, is the essential mechanism by which all living things reproduce. Dividing cells must coordinate their growth. A complex network of regulatory proteins trigger the different events of the cycle.
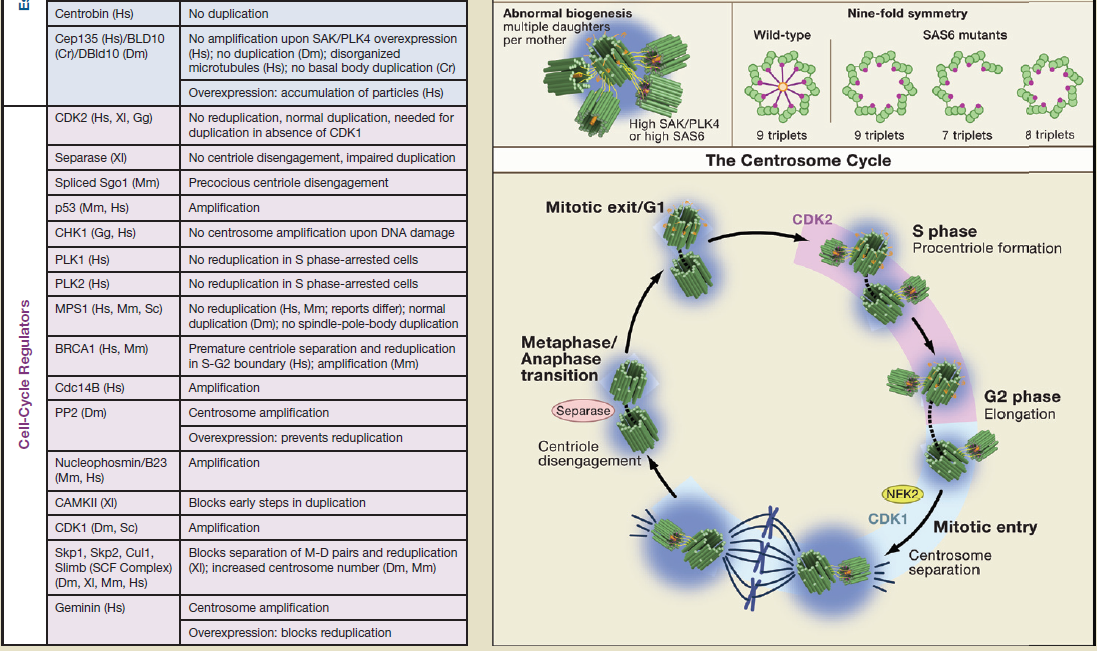
During the cell cylce, eighteen different regulators are required, which order and coordinate the process. Each of these regulators are absolutely essential. If one is missing, the cell cycle is not completed and, the cell cannot duplicate. Any of these regulators have only use if fully integrated in the process. They have no use or function by themself. This makes replication a irreducible , interdependent process.
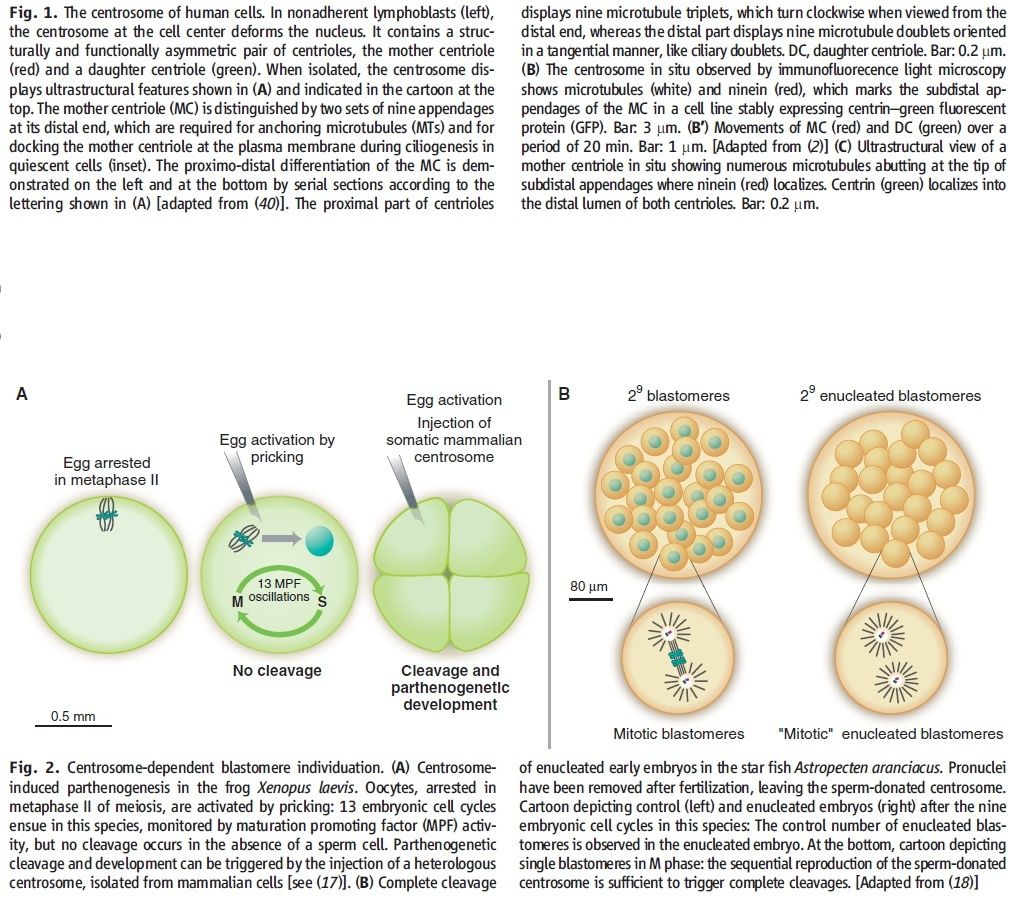
Centrosomes play a key role in organizing the microtubule network of the cell, most notably the mitotic spindle during cell division .
The choreography of microtubules, centrosomes and chromosomes during mitosis and meiosis is beautifully designed, and uses finely regulated and synchronized movements.
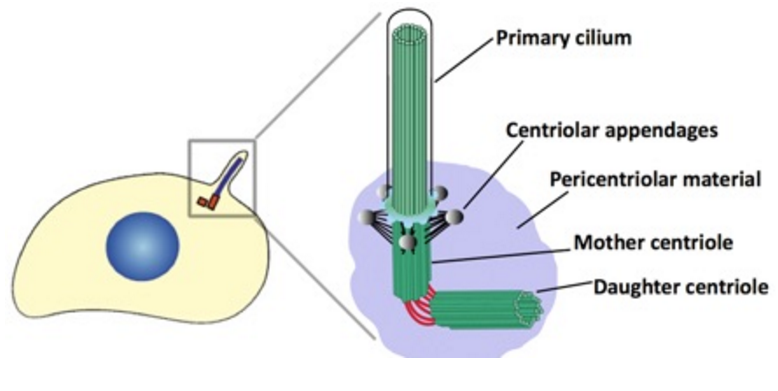
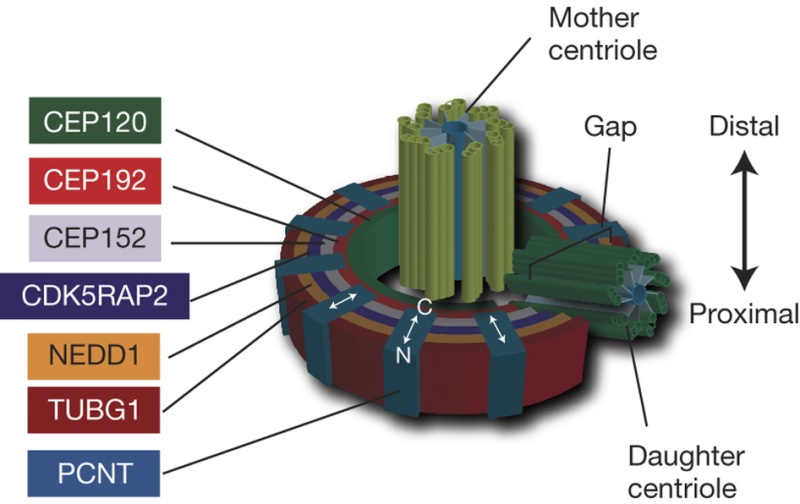
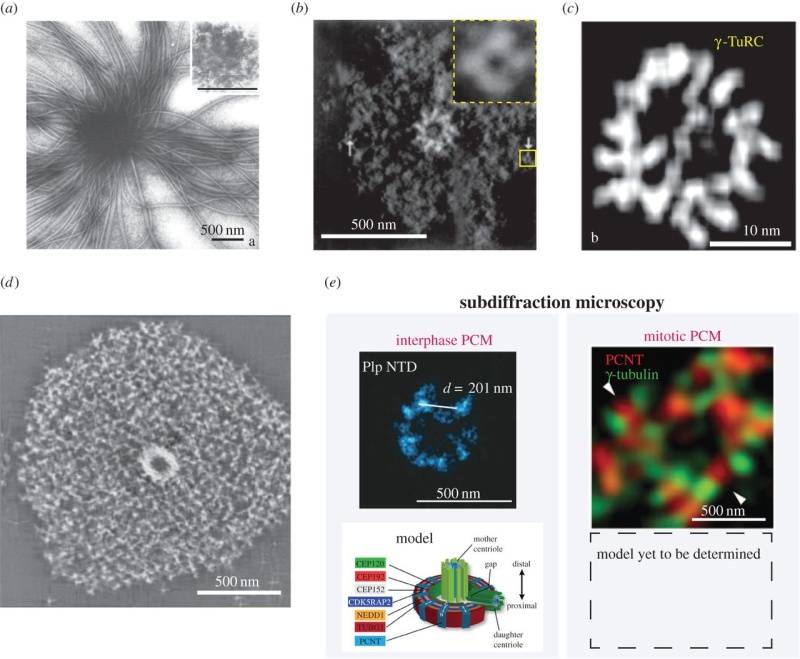
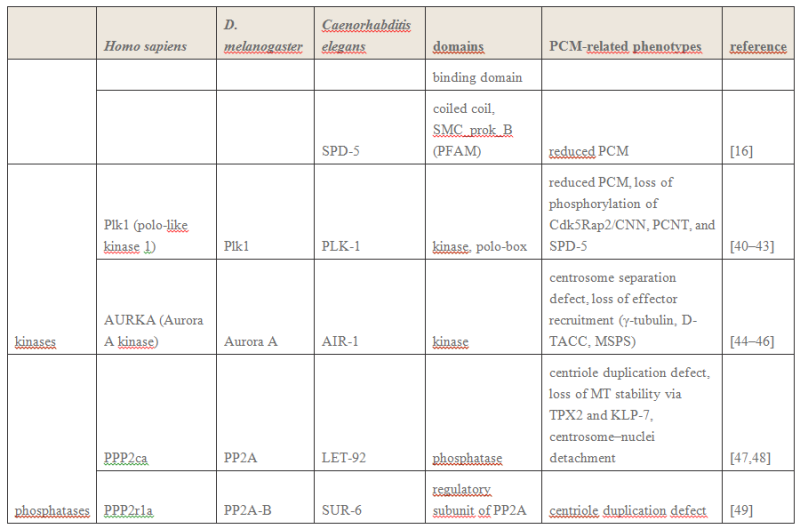
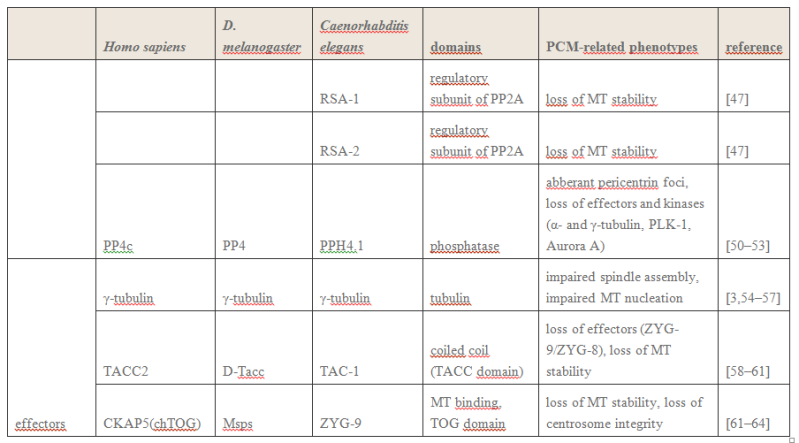
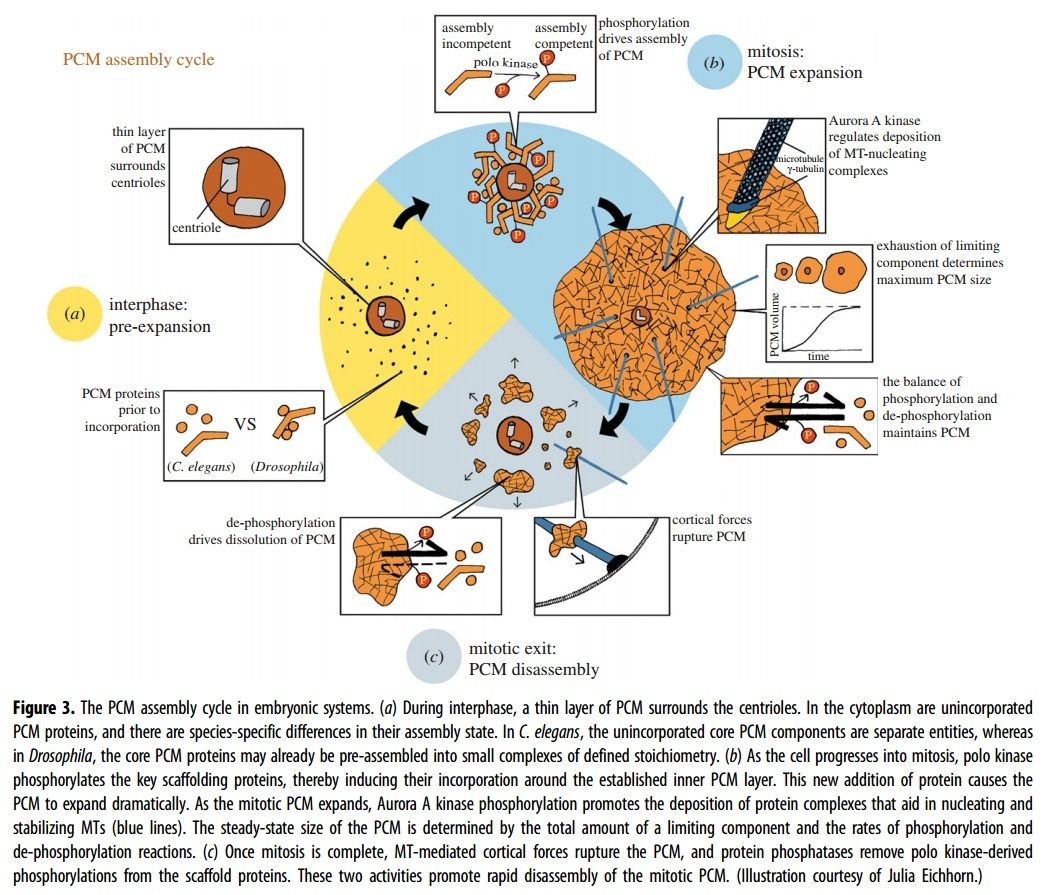
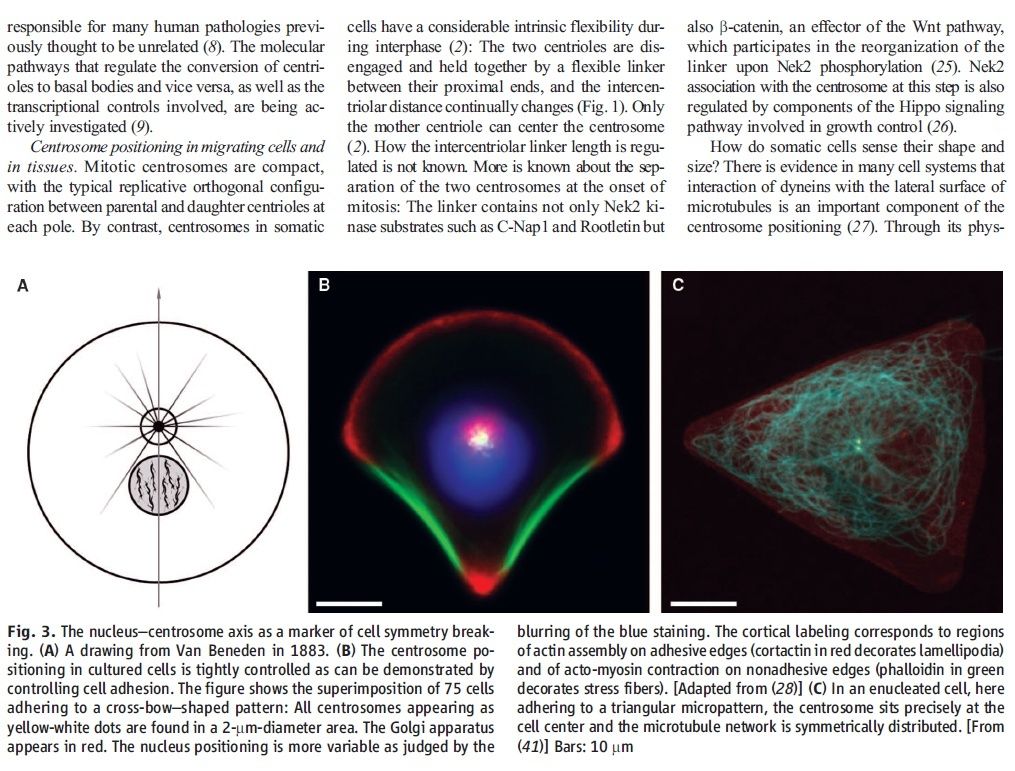
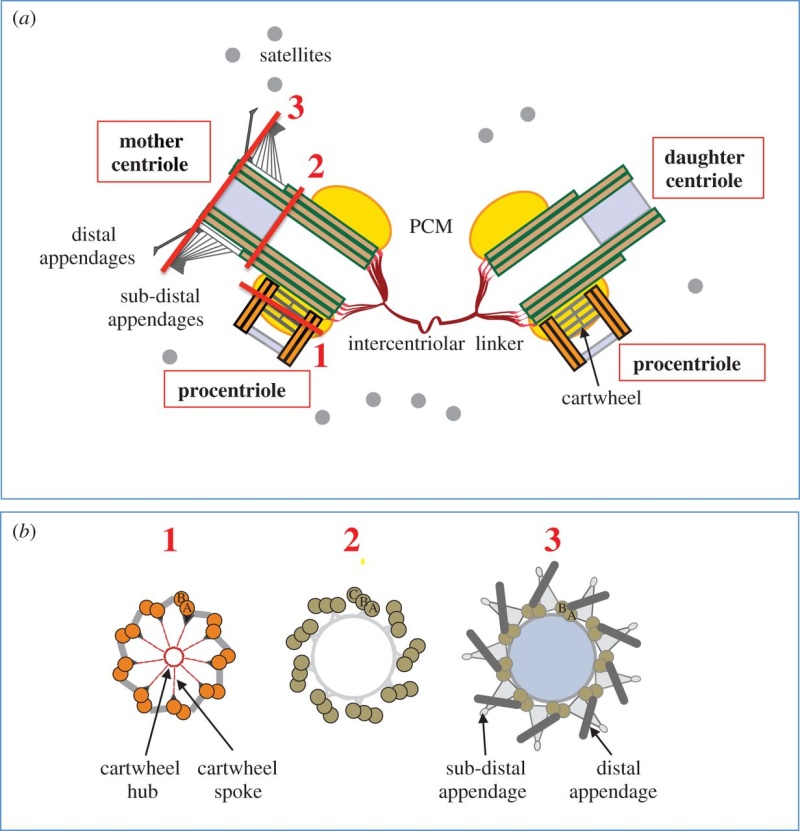
The centrosome is a structure, consisting of a pair of cylindrical microtubule-based organelles called centrioles , embedded in an amorphous network of proteins known collectively as Pericentriolar Material (PCM). Microtubules (MTs) originate from the PCM. The PCM comprises a porous structural scaffold onto which γ-tubulin and other soluble components from the cytoplasm are loaded. Centrosome growth is an aggregation process of a condensed phase of PCM components, which segregate from the cytosol. The aggregation process leads to a centrosome phase that coexists with the cytosol and does rearrange internally. This implies that the centrosome phase is viscoelastic, such that on long timescales it behaves as a liquid-like material.
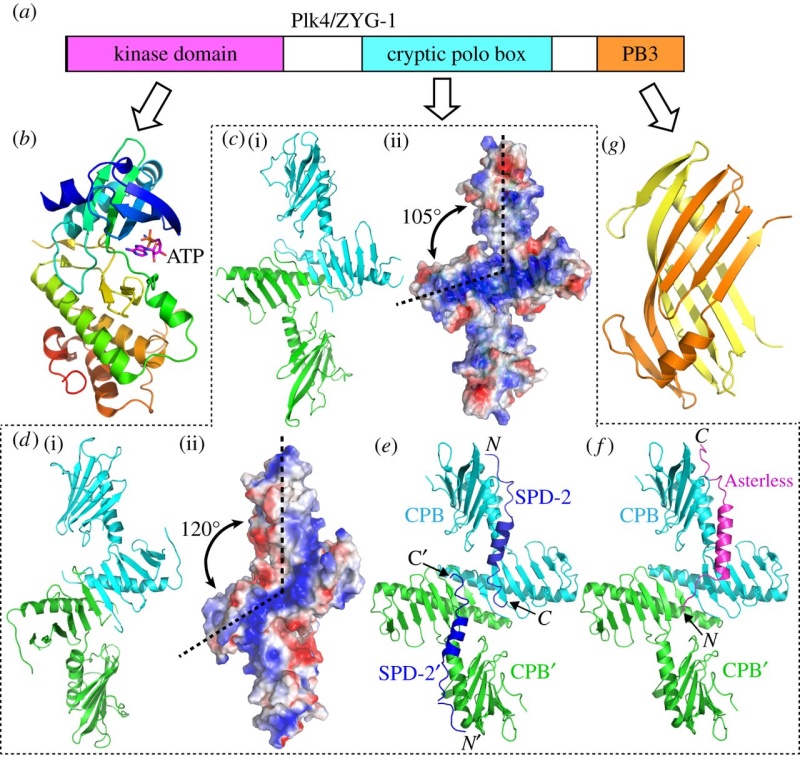
Cep192 is a pericentriolar protein that accumulates at centrosomes during mitosis and is required for PCM recruitment, centriole duplication, microtubule nucleation, and centrosome maturation.
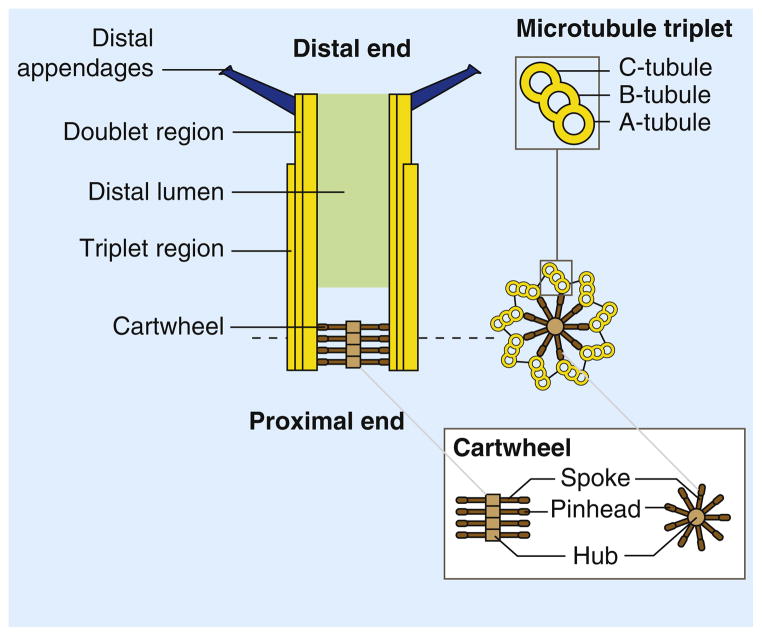
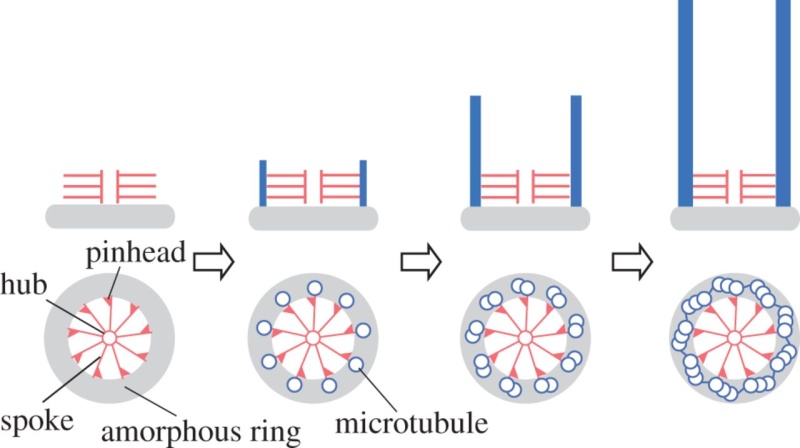
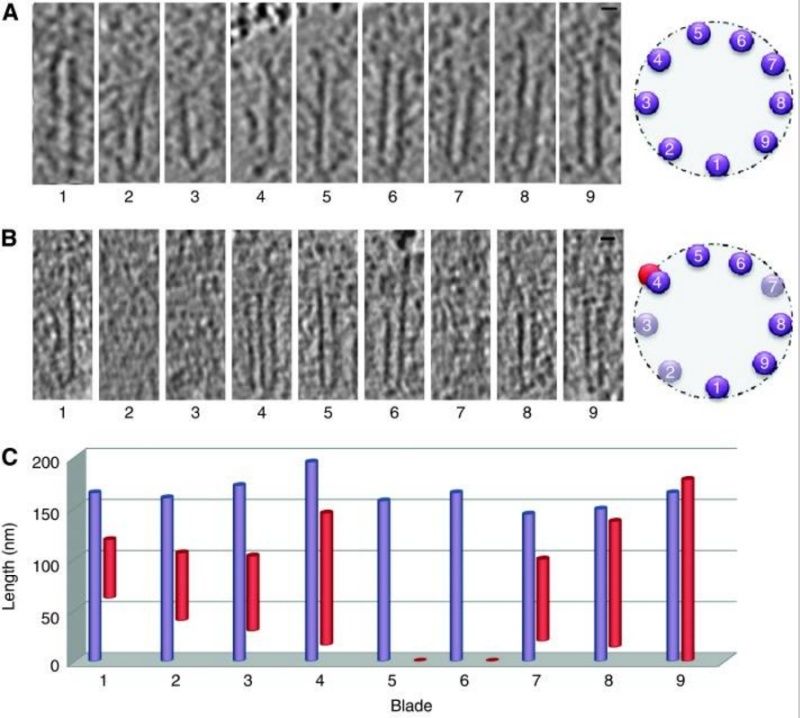
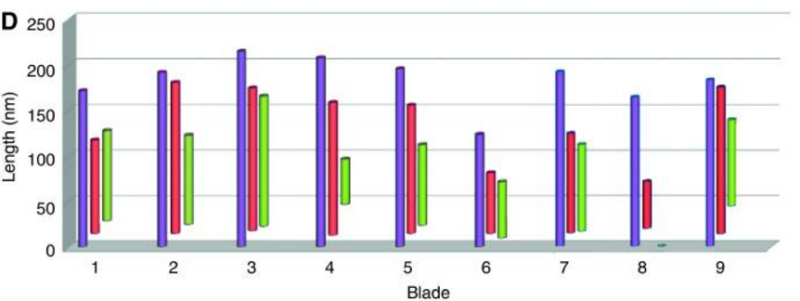
Centrioles are among the most beautiful of biological structures. How their highly conserved nine-fold symmetry is generated is a question that has intrigued cell biologists for decades.
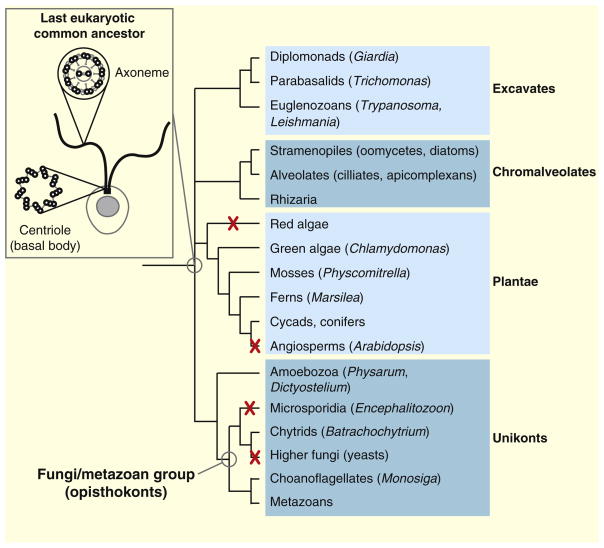
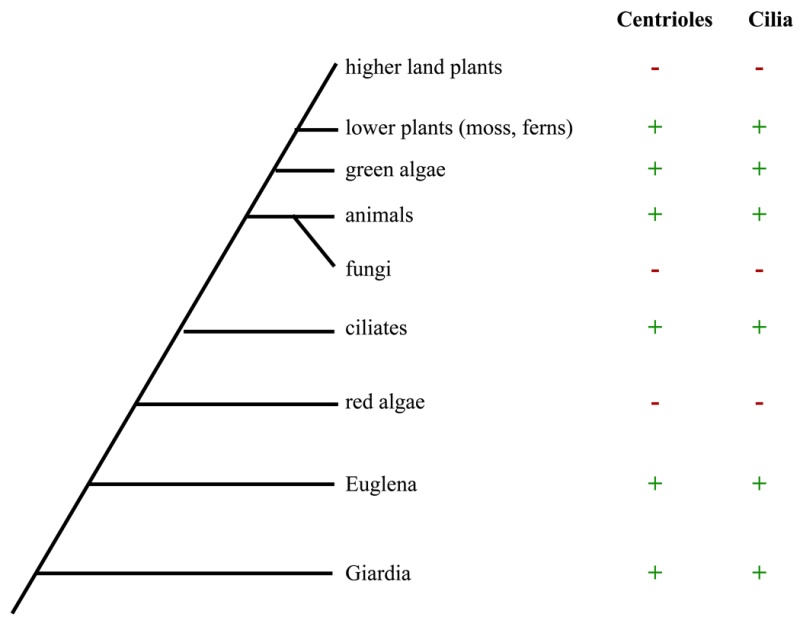
Centrioles are present in all eukaryotic species that form cilia and flagella, but are absent from higher plants and higher fungi which do not have cilia.
It seems likely that they have the primary purpose of growing cilia and flagella, which are important sensory and motile organelles found in almost all cells of the human body. These organelles have many important functions in cells, and their dysfunction has been linked to a plethora of human pathologies, ranging from cancer to microcephaly to obesity. Great progress has been made recently in understanding how these proteins interact and how these interactions are regulated to ensure that a new centriole is only formed at the right place and at the right time.
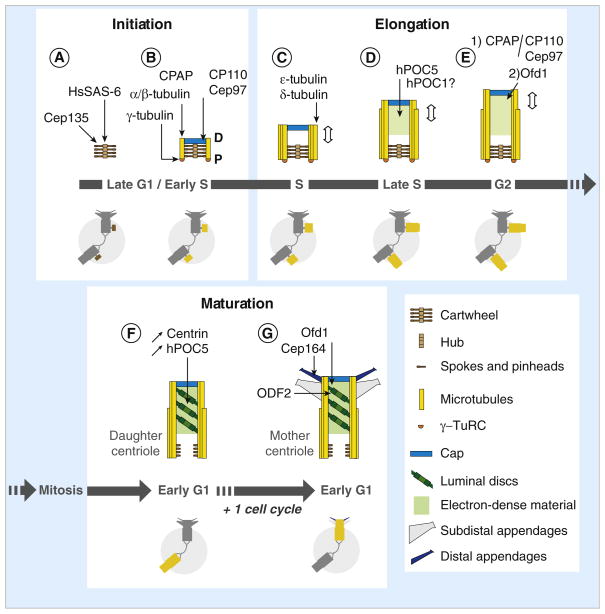
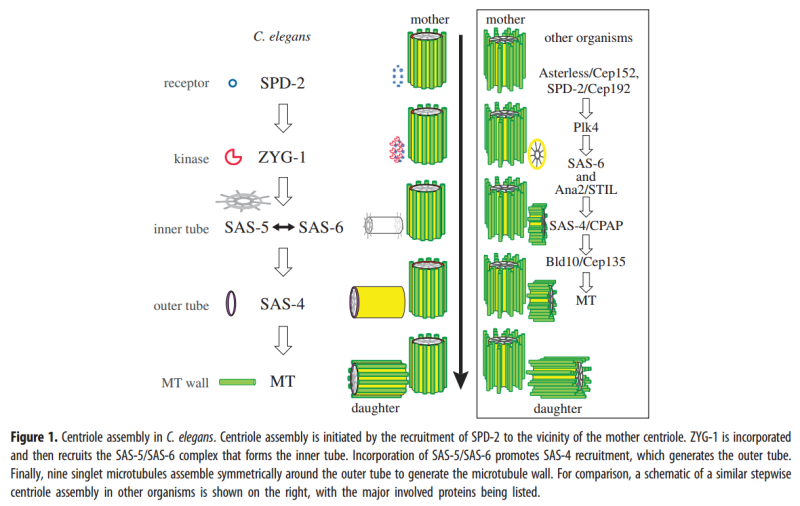
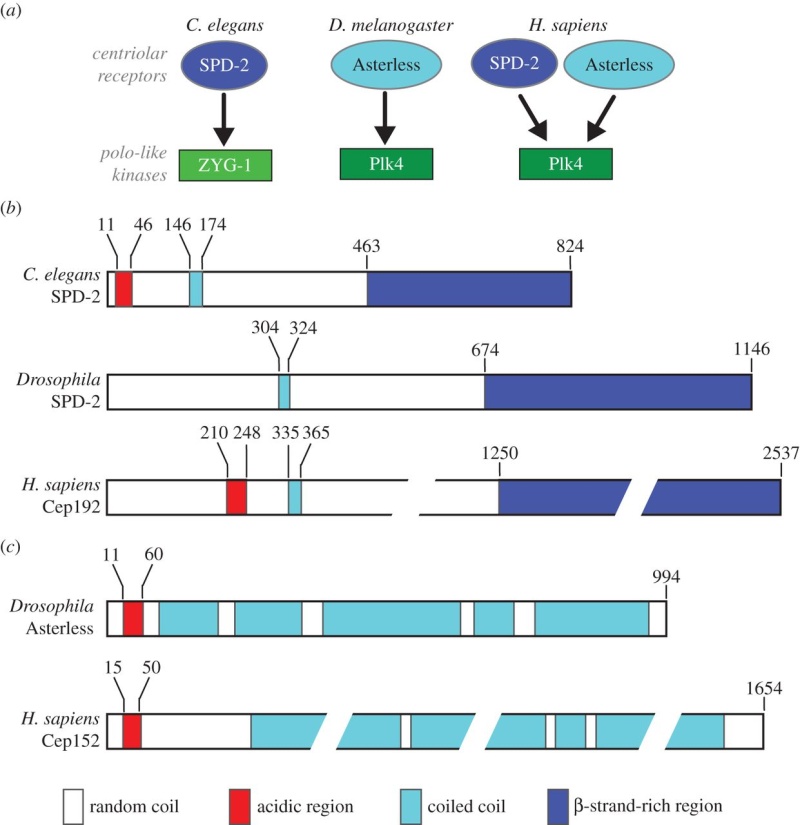
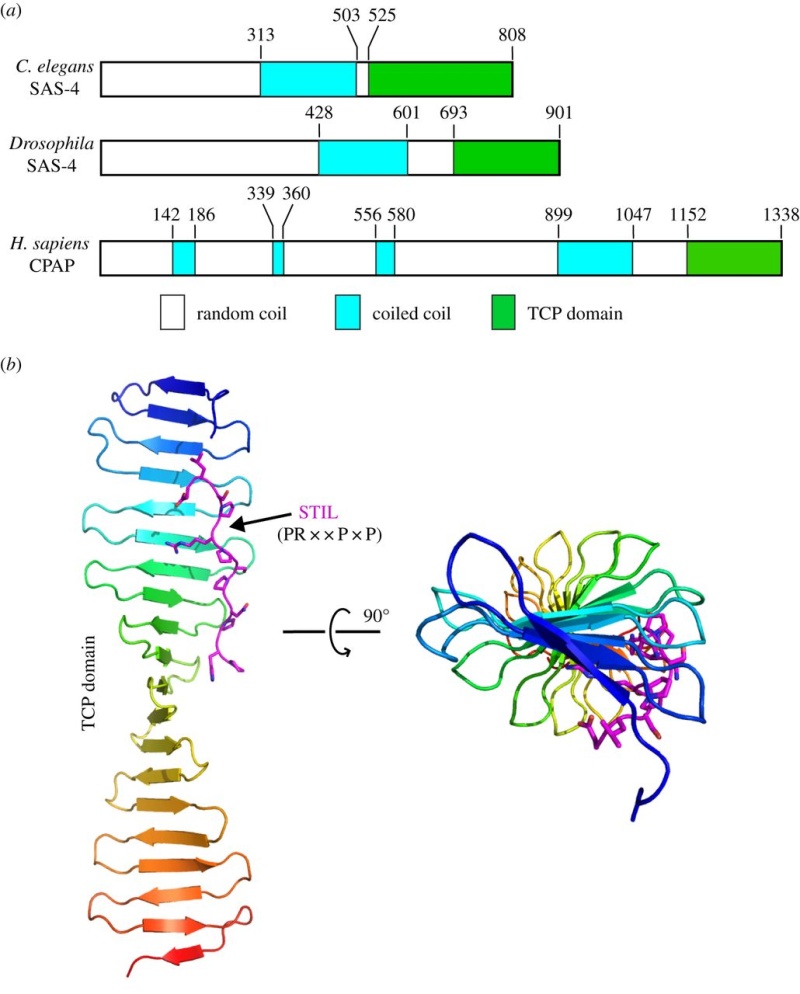
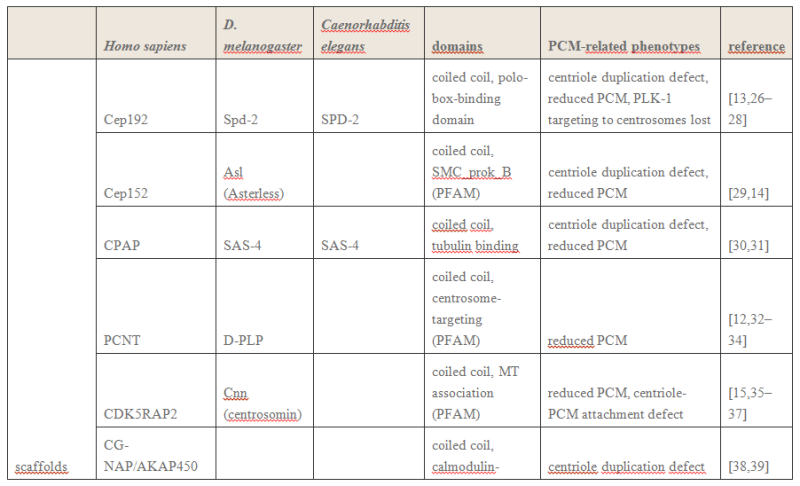
Centriole biogenesis requires thirteen essential molecules. If any of these molecules is missing, centrioles cannot be made.
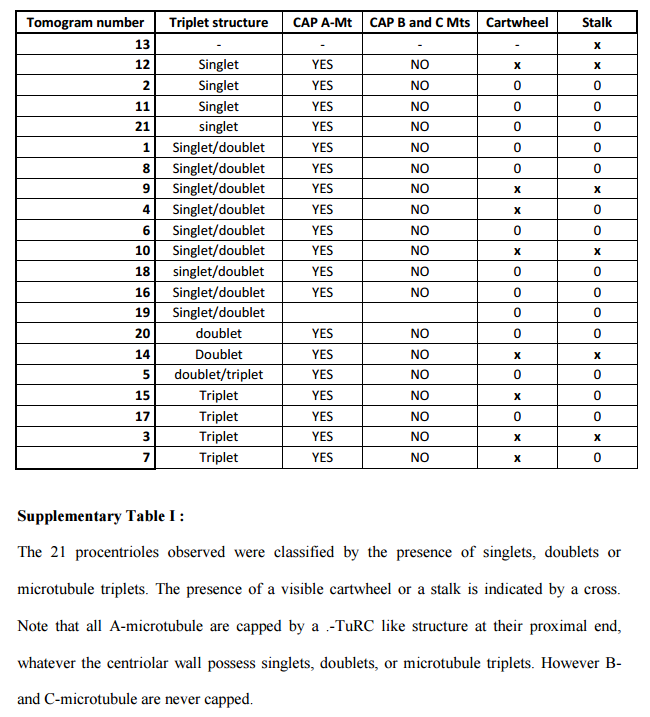
Centriole assembly is also tightly regulated and abnormalities in this process can lead to developmental defects and cancer. Initiation of centriole duplication is under tight regulation to ensure the control of centriole number. Presumably in centriole initiation, there is some form of cooperativity or positive feedback that results in asymmetric accumulation of the relevant proteins in a symmetric background.
So we have not only the requirement of eighteen proteins required for cell cycle regulation, but also thirteen essential molecules for centriole biogenesis, which by itself is also tighthly regulated, requiring positive feedback.
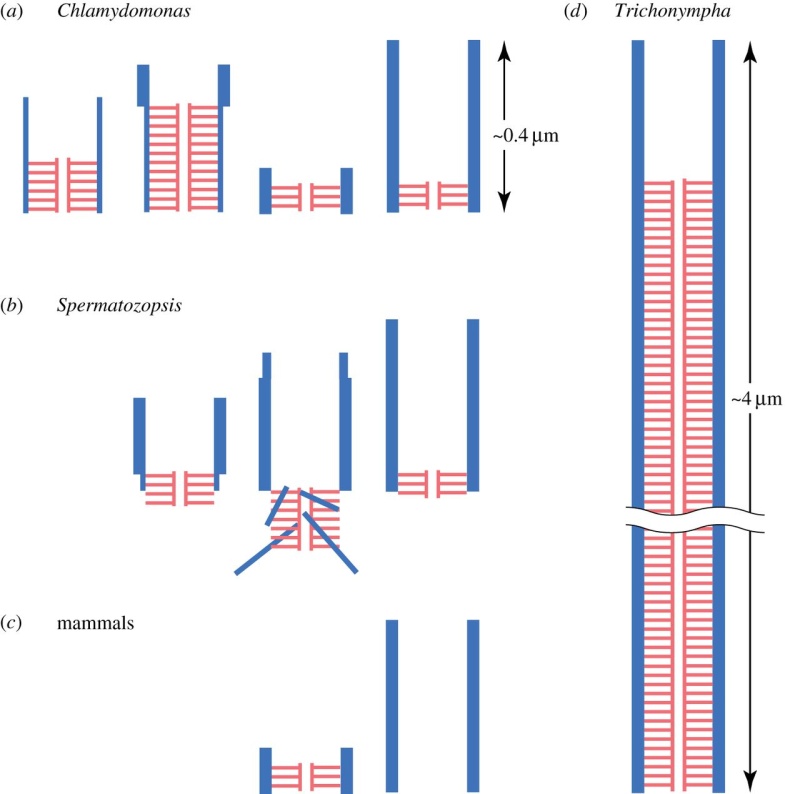
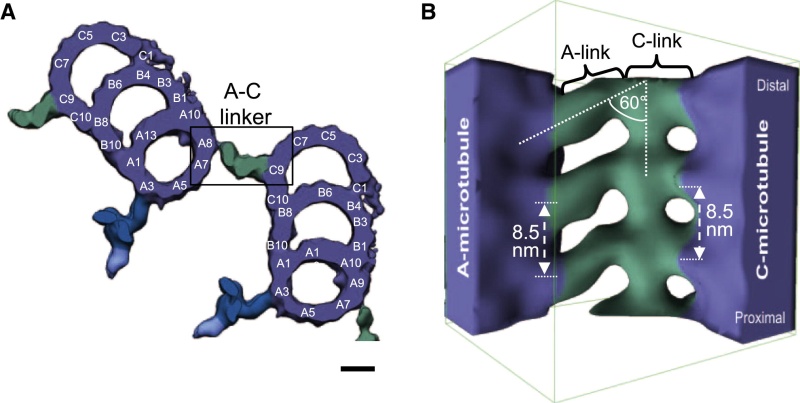
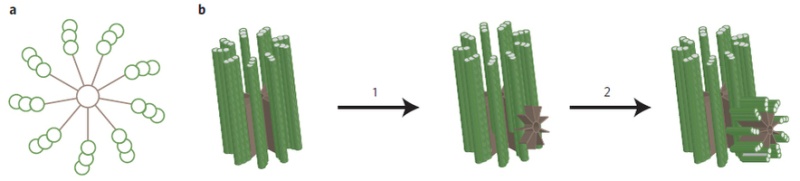
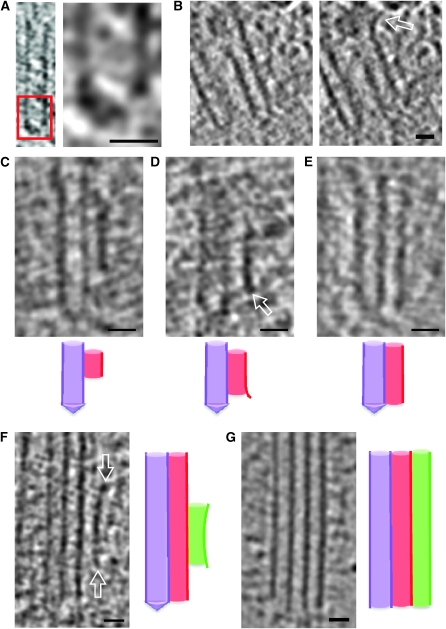
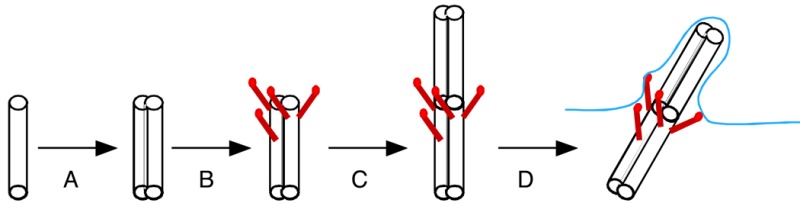
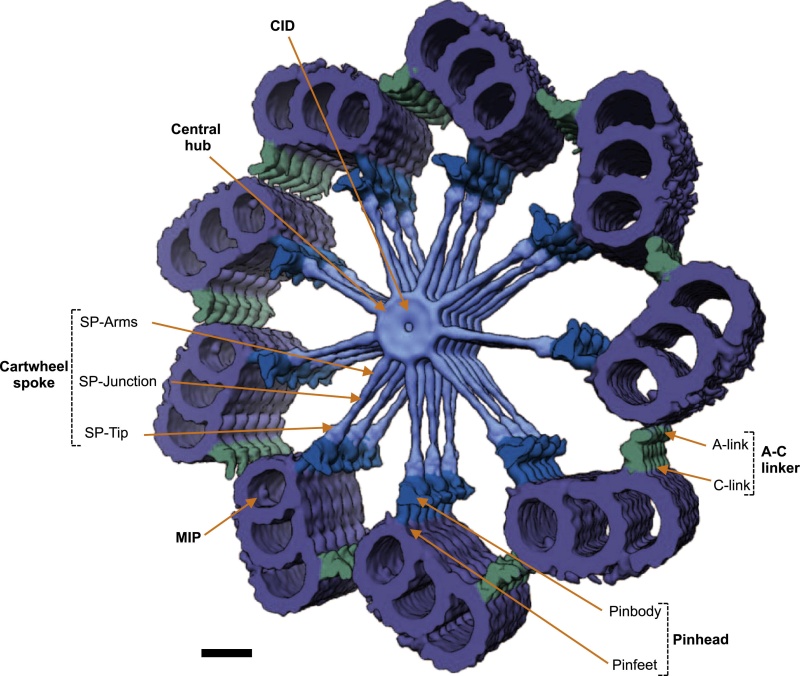
It appears at the initial stage of the centriole assembly process as the first ninefold symmetrical structure. The cartwheel was first described more than 50 years ago, but it is only recently that its pivotal role in establishing the ninefold symmetry of the centriole was demonstrated. This is a highly ordered structure that really stands out from the background. Constructed of rod-like microtubules, most centrioles have a nine-fold pattern, nine triplets or doublets evenly spaced at the rim, giving it a "cartwheel" appearance in cross-section.
The comparison to a human made cartwheel is evident, and so that it is intelligently designed. Obviously, the question arises, how could all this emerge gradually ?
Another amazing fact is that Electromagnetics play an important role in cell functioning and especially in cell duplication and division (mitosis).
Recent development in the field of quantum biology highlights that the intracellular electromagnetic field (EMF) of microtubules plays an important role in many fundamental cellular processes such as mitosis. It is an intriguing hypothesis that centrosome functions as molecular dynamo to generate electric flow over the microtubules, leading to the electric excitation of microtubule EMF that is required for spindle body microtubule self-assembly. With the help of motors proteins within the centrosome, centrosome transforms the energy from ATP into intracellular EMF in the living cell that shapes the functions of microtubules. There will be a general impact for the cell biology field to understand the mechanistic function of centrosome for the first time in correlation with its structural features.
The electromagnetic property of microtubule has been reported with both computation modelling and experimental evidences.
To transform the chemical energy in ATP into electric magnetic field within the living cell, cell needs to have a molecular dynamo to transform the mechanistic movement of protein complexes to directional movements of intracellular electrons, leading to the electric excitation of the spindle body microtubules as well as the M phase chromosomes, which is essential for mitosis
Taken together the longitudinal, or axial, vibration of the 13 filaments of an MT and then the 27 MTs making up the centriole barrel produce the electromagnetic field surrounding the centriole . Interestingly, this field is also found to be ferromagnetic. Also of interest, the fundamental vibration frequency of an MT filament is approximately 465 MHz, although this frequency is continually changing due to the ongoing length changing of the filaments. The electropolarity of the centrioles enables them to exert forces at a distance—that is, forces without physical contact.
"All this cannot [but] leave us astounded[; it] indicates the requirement of for[e]sight to produce all these ingenius[ingenious] mechanisms,and subsequently[consequently] a[n] intelligent designer.
Yuan Tian: Nine-fold symmetry of centriole: The joint efforts of its core proteins 07 January 2022
https://onlinelibrary.wiley.com/doi/abs/10.1002/bies.202100262
https://reasonandscience.catsboard.com/t2090-centriole-biogenesis-and-the-duplication-cycle-amazing-evidence-of-design
https://reasonandscience.catsboard.com/t2090-centriole-biogenesis-and-the-duplication-cycle-amazing-evidence-of-design
The duplication of eukaryotic cells is a all fine-tuned biochemical processes that depends on the precise structural arrangement of the cellular components. Mitotic cell division is the most fundamental task of all living cells. Cells have intricate and tightly regulated machinery to ensure that mitosis occurs with appropriate frequency and high fidelity.
The only way to make a new cell is to duplicate a cell that already exists. A cell reproduces by performing an orderly sequence of events in which it duplicates its contents and then divides in two. This cycle of duplication and division, known as the cell cycle, is the essential mechanism by which all living things reproduce. Dividing cells must coordinate their growth. A complex network of regulatory proteins trigger the different events of the cycle.
During the cell cylce, eighteen different regulators are required, which order and coordinate the process. Each of these regulators are absolutely essential. If one is missing, the cell cycle is not completed and, the cell cannot duplicate. Any of these regulators have only use if fully integrated in the process. They have no use or function by themself. This makes replication a irreducible , interdependent process.
Centrosomes play a key role in organizing the microtubule network of the cell, most notably the mitotic spindle during cell division .
The choreography of microtubules, centrosomes and chromosomes during mitosis and meiosis is beautifully designed, and uses finely regulated and synchronized movements.
The centrosome is a structure, consisting of a pair of cylindrical microtubule-based organelles called centrioles , embedded in an amorphous network of proteins known collectively as Pericentriolar Material (PCM). Microtubules (MTs) originate from the PCM. The PCM comprises a porous structural scaffold onto which γ-tubulin and other soluble components from the cytoplasm are loaded. Centrosome growth is an aggregation process of a condensed phase of PCM components, which segregate from the cytosol. The aggregation process leads to a centrosome phase that coexists with the cytosol and does rearrange internally. This implies that the centrosome phase is viscoelastic, such that on long timescales it behaves as a liquid-like material.
Cep192 is a pericentriolar protein that accumulates at centrosomes during mitosis and is required for PCM recruitment, centriole duplication, microtubule nucleation, and centrosome maturation.
Centrioles are among the most beautiful of biological structures. How their highly conserved nine-fold symmetry is generated is a question that has intrigued cell biologists for decades.
Centrioles are present in all eukaryotic species that form cilia and flagella, but are absent from higher plants and higher fungi which do not have cilia.
It seems likely that they have the primary purpose of growing cilia and flagella, which are important sensory and motile organelles found in almost all cells of the human body. These organelles have many important functions in cells, and their dysfunction has been linked to a plethora of human pathologies, ranging from cancer to microcephaly to obesity. Great progress has been made recently in understanding how these proteins interact and how these interactions are regulated to ensure that a new centriole is only formed at the right place and at the right time.
Centriole biogenesis requires thirteen essential molecules. If any of these molecules is missing, centrioles cannot be made.
Centriole assembly is also tightly regulated and abnormalities in this process can lead to developmental defects and cancer. Initiation of centriole duplication is under tight regulation to ensure the control of centriole number. Presumably in centriole initiation, there is some form of cooperativity or positive feedback that results in asymmetric accumulation of the relevant proteins in a symmetric background.
So we have not only the requirement of eighteen proteins required for cell cycle regulation, but also thirteen essential molecules for centriole biogenesis, which by itself is also tighthly regulated, requiring positive feedback.
It appears at the initial stage of the centriole assembly process as the first ninefold symmetrical structure. The cartwheel was first described more than 50 years ago, but it is only recently that its pivotal role in establishing the ninefold symmetry of the centriole was demonstrated. This is a highly ordered structure that really stands out from the background. Constructed of rod-like microtubules, most centrioles have a nine-fold pattern, nine triplets or doublets evenly spaced at the rim, giving it a "cartwheel" appearance in cross-section.
The comparison to a human made cartwheel is evident, and so that it is intelligently designed. Obviously, the question arises, how could all this emerge gradually ?
Another amazing fact is that Electromagnetics play an important role in cell functioning and especially in cell duplication and division (mitosis).
Recent development in the field of quantum biology highlights that the intracellular electromagnetic field (EMF) of microtubules plays an important role in many fundamental cellular processes such as mitosis. It is an intriguing hypothesis that centrosome functions as molecular dynamo to generate electric flow over the microtubules, leading to the electric excitation of microtubule EMF that is required for spindle body microtubule self-assembly. With the help of motors proteins within the centrosome, centrosome transforms the energy from ATP into intracellular EMF in the living cell that shapes the functions of microtubules. There will be a general impact for the cell biology field to understand the mechanistic function of centrosome for the first time in correlation with its structural features.

The electromagnetic property of microtubule has been reported with both computation modelling and experimental evidences.
To transform the chemical energy in ATP into electric magnetic field within the living cell, cell needs to have a molecular dynamo to transform the mechanistic movement of protein complexes to directional movements of intracellular electrons, leading to the electric excitation of the spindle body microtubules as well as the M phase chromosomes, which is essential for mitosis
Taken together the longitudinal, or axial, vibration of the 13 filaments of an MT and then the 27 MTs making up the centriole barrel produce the electromagnetic field surrounding the centriole . Interestingly, this field is also found to be ferromagnetic. Also of interest, the fundamental vibration frequency of an MT filament is approximately 465 MHz, although this frequency is continually changing due to the ongoing length changing of the filaments. The electropolarity of the centrioles enables them to exert forces at a distance—that is, forces without physical contact.
"All this cannot [but] leave us astounded[; it] indicates the requirement of for[e]sight to produce all these ingenius[ingenious] mechanisms,and subsequently[consequently] a[n] intelligent designer.
Yuan Tian: Nine-fold symmetry of centriole: The joint efforts of its core proteins 07 January 2022
https://onlinelibrary.wiley.com/doi/abs/10.1002/bies.202100262
https://reasonandscience.catsboard.com/t2090-centriole-biogenesis-and-the-duplication-cycle-amazing-evidence-of-design
Last edited by Otangelo on Sat 18 Jun 2022 - 19:26; edited 22 times in total